Сезонные изменения состава населения и численности рыб
В западно-камчатских реках массовый выход из грунта и скат молоди тихоокеанских
лососей и гольцов приурочены к подъему уровня воды в мае-июне (Павлов и др., 2006; Pavlov
et al., 2005). С наступлением неустойчивой летней межени в июле-сентябре максимальное
количество молоди и взрослых жилых рыб включаются в состав относительно стабильных
территориальных агрегаций (Панов, 1983; Живоглядов, 2004; Root, 1967). Значительное пере-
распределение рыб в небольших лососевых водотоках в летние месяцы возможно лишь в
случаях сильных паводков или пересыхания русел (Богатов, 1994; McCart, 1967; Scarlett,
Cederholm, 1984; Connor, 2002). Обострение пространственной и пищевой конкуренции в этот
период (Леванидов, 1981) приводит к расхождению разных размерно-видовых групп рыб по
разным зонам и биотопам (Леман и др., 2005; Ермакова и др., 2006).
Имеющиеся сведения о зимовке молоди и взрослых жилых рыб в малых лососевых
водотоках весьма противоречивы. По одним данным большая часть рыб в конце осени
мигрирует вниз по течению на зимовальные ямы (Никольский, 1953; Христофоров, 1982;
Живоглядов, 2004), по другим сведениям большинство рыб зимует на участках летне-осеннего
нагула (Сunjak, 1996; Huusko et al., 2007). Часто поведение лососевых рыб зимой определяется
условиями конкретного года (Pacific salmon…, 1995; Enders et al., 2007). Известно, что после
понижения температуры воды до 3 ºС скорость метаболизма зимующей в руслах молоди и
взрослых жилых лососей снижается на порядок, что приводит к исчезновению конкуренции
за пищу и пространство (Chapman, 1966; Huusko et al., 2007). Выбор местообитания зимой опре-
деляется, прежде всего, необходимостью минимизировать затраты энергии и при этом
защититься от промерзания или пересыхания местообитания, недостатка кислорода и т.п.
(Riehle, Griffith, 1993; Сunjak, 1996). Доступ к пище при температурах, близких к замерзанию,
обычно не является приоритетом при выборе укрытия (Сunjak, 1996; Enders et al., 2007). Замет-
ная пищевая активность у молоди большинства видов лососей начинает проявляться при
температуре воды, близкой к 4—5 ºС (Griffith, Smith, 1993), т.е. в малых западно-камчатских
реках активный поиск пищи молодью начинается не ранее конца мая (см. рис. 2.2).
6.1.1. Порожисто-водопадные русла
Нереста рыб на порожистых руслах не происходит, половозрелые проходные и жилые
лососи в ступенчатые горные истоки р. Начилова не поднимаются. Летнее население образо-
вано молодью камчатской мальмы (табл. 6.1).
Половодье (май-июнь). Верховья реки вскрываются ото льда в конце мая, когда ниже по
течению половодье уже идет на убыль. Подъем уровня на порожистых руслах кратковременный
и продолжается 5—10 дней. Рыбы в это время в зоне истоков нет.
Меженно-паводочный период (июль-октябрь). Молодь заходит в порожистые водотоки,
начиная с июня. За время июльских исследований в зоне было поймано 13 двухлеток мальмы,
92
в сентябре — 3 двухлетки, молоди прочих видов не обнаружено. В июле 2005—2006 гг. мальма
ловилась в водоворотных зонах эрозионных ям под гребнями порогов, у берегов рыбы заме-
чено не было. Средняя июльская плотность и биомасса рыбного населения минимальна —
оцененные показатели не достигали 0,1 экз. и г на м2 дна соответственно.
Устойчивая межень (ноябрь-апрель). Начиная с ноября (устойчивая отрицательная
температура воздуха), эрозионные ямы под порогами закрываются льдом и снегом, течение
воды прекращается. По-видимому, рыбы скатываются из истоков в октябре-ноябре. На протя-
жении зимы молодь и взрослые особи в порожистых водотоках отсутствуют.
6.1.2. Горные русла без перекатов
В верхнее течение на нерест поднимаются потамодромная мальма, кижуч раннего и
позднего нереста, а также проходная сима. Круглый год в районе нерестилищ держатся
карликовые самцы мальмы и симы.
Круглый год на горных руслах без перекатов также держится молодь лососевых. За
время сезонных исследований здесь были пойманы личинки, мальки и пестрятки мальмы,
кижуча и симы (табл. 6.1). Смолтификация этих видов происходит ниже по течению в зоне
равнинных русел с перекатами.
Половодье (май-июнь). Подъем уровня воды на врезанных руслах не приводит к значи-
тельному увеличению акватории, новые местообитания не образуются. Река превращается в
стремительный бурный поток. Двух- и трехгодовики мальмы, а также карликовые половозрелые
самцы мальмы и симы (возраст 3·) в начале мая укрываются за крупными камнями на глайдах и
в редких периферических водоемах. Личинки мальмы, которые только начинают выходить из
бугров, укрываются в межвалунном пространстве в прибрежье и в первой декаде мая почти не
мигрируют с нерестилищ. Личинки и мальки кижуча (0·) выходят в приповерхностные слои
воды, подхватываются потоком и уносятся вниз по течению. В время исследований распреде-
ление рыб по акватории было крайне неравномерным — весной 2007 г. за валунами на отмелях
отмечались скопления плотностью до 15—20 экз./м2 дна, на потоке рыбы почти не было.
Меженно-паводочный период (июль-октябрь). Молодь мальмы в возрасте 1+ и 2+, а
также карликовые самцы мальмы и симы в июле распределяются по эрозионным ямам
и локальным укрытиям на глайдах, которые представляют собой участки замедленного течения
за крупными одиночными камнями и древесными заломами. Течение в таких укрытиях почти
отсутствует (часто оно обратное), освещенность снижена. Подросшие мальки мальмы рассе-
ляются по прибрежным отмелям вблизи нерестилищ, в момент опасности они укрываются под
камнями. Пополнение сеголеток мальмы на отмелях продолжается до середины июля за счет
растянутого выхода из бугров. Выход личинок кижуча завершается в конце июля, сеголетки за-
нимают периферические биотопы, где нередко образуют смешанные стайки с мальмой. На
протяжении всего лета молодь кижуча откочевывает из зоны — вышедшие на глайды мальки не
способны противостоять потоку, подхватываются им и уносятся вниз по течению. Известно, что
интенсивность такой пассивной миграции значительно возрастает во время паводков (Giannico,
Healey, 1998). К октябрю молодь кижуча полностью покидает зону. Большая часть сеголеток
симы, выходящих из грунта в июне, сносится течением вниз. Оставшаяся часть молоди рассе-
ляется по отмелям и притокам, в т.ч. по устьям овражных водотоков. Здесь сима держится до
зимы.
В июле 2006 г. средняя плотность обитания молоди и жилых лососей на прибрежных
отмелях, в заводях и эрозионных ямах была выше 1 экз./м2. В верхней части зоны (станция Н2)
единовременно было заселено не более 40 % укрытий всех типов, плотность в них достигала
3 экз./м2 (в среднем 1,5 экз./м2). Ниже по течению (Н3) в это же время было заселено 50 %
укрытий, плотность в них достигала 7 экз./м² (в среднем 1,6 экз./м²). Пороги и большая часть
открытых глайдов оставались малообитаемы и труднопреодолимы для молоди (табл. 6.2).
Средняя июльская плотность обитания молоди и жилых форм лососей повышалась вниз
по течению зоны с 0,1 (Н2) до 0,3 (Н3) экз./м2 участка реки, включающего все характерные типы
русловых и периферических местообитаний. Средняя биомасса рыб увеличивалась с 0,2 до
3,25 г/м2. Средняя масса 1 экз. лососевых рыб в июле колебалась от 1,9 (Н2) до 10,8 (Н3) г.
С конца июля на неразветвленные горные русла поднимаются производители потамо-
дромной мальмы и проходной симы, которые занимают глубокие русловые ямы. Совместный
летний нерест проходной формы и карликовых самцов симы происходит на галечниковых уча-
стках глайдов перед порогами. Мальма нерестится в сентябре-октябре на глайдах и в устьях
овражных притоков. В октябре-ноябре жилые производители мальмы покидают зону. Нерест
кижуча начинается с конца сентября на глайдах, прибрежных отмелях и в эрозионных ямах.
93
Таблица 6.1. Состав сезонных ассамблей молоди и взрослых жилых рыб в бассейне р. Начилова в 2006—2007 гг. (%)
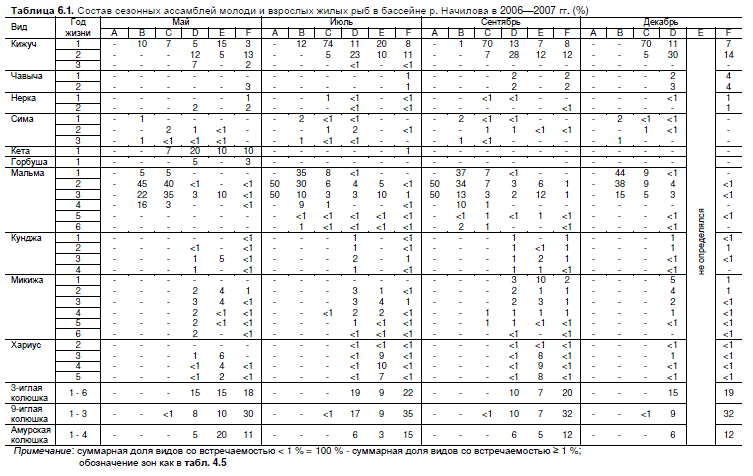
Примечание: суммарная доля видов со встречаемостью < 1 % = 100 % - суммарная доля видов со встречаемостью ≥ 1 %;
обозначение зон как в табл. 4.5
94
Таблица 6.2. Характеристика ассамблеи молоди и взрослых жилых рыб на горных руслах
без перекатов в июле
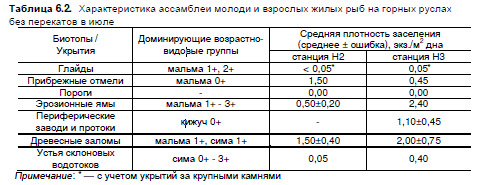
Примечание: * — с учетом укрытий за крупными камнями
Устойчивая межень (ноябрь-апрель). Зимой 2006 г. на неразветвленных горных руслах
ловилась молодь мальмы и симы, а также карликовые самцы симы. В начале декабря при тем-
пературе воды 3—4 ºС сеголетки покидают периферические местообитания и прибрежные
отмели, где они удерживали участки дна площадью по 0,1—0,2 м2, и перемещаются на русло-
вые глайды под ледяные забереги. Здесь молодь укрывается в межвалунном пространстве и
большую часть времени не проявляет двигательной активности. Большая часть двух- и трехле-
ток, как и осенью, держится в центральной части русла за валунами и в глубоких ямах на
потоке. После замерзания реки рыбы всех размеров перемещаются на участки средней глу-
бины на границе льда и потока. Большая часть мелких рыб держится под камнями, крупные
особи предпочитают положение у дна за валунами или обломками скал. Большую часть вре-
мени двух- и трехлетки неподвижны и занимают позицию в контакте с дном в 15—30 см позади
рассекающего поток объекта (наши наблюдения; Erkinaro et al., 1994; Enders et al., 2007).
Нередко рыбы используют в качестве укрытий выступы дна, по высоте сопоставимые с высотой
их собственного тела. Во время исследований молодь была равномерно распределена по
открытым участкам русла с плотностью 0,15 экз./м2.
Обычно ледовая обстановка не меняется в зоне горных русел без перекатов до апреля,
донный лед не образуется. В этих стабильных условиях молодь лососей должна вести себя
однотипно в течение всей зимы и далеко не мигрировать (Enders et al., 2007). В апреле перед
половодьем из гнезд начинает выходить кижуч, который скапливается на отмелях под забере-
гами.
Смотрите также






