Выделение биотопов малых лососевых водотоков
Русло любого типа и морфологического строения состоит из набора повторяющихся
систем русловых форм, или макробиотопов. Всего в малых лососевых водотоках Западной
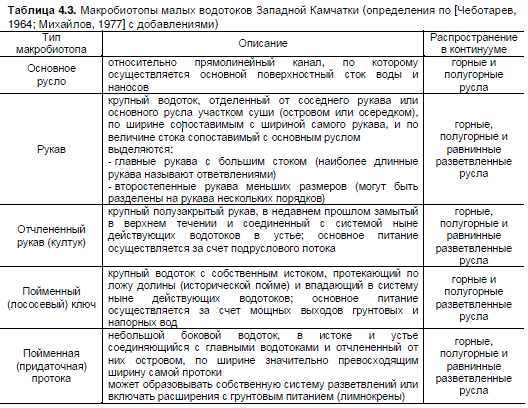
Камчатки выделены 8 типов макробиотопов, к которым относятся основное русло, излучины,
водотоки разветвлений, а также их трансформированные элементы — старицы, отшнурован-
ные протоки и т.д. (табл. 4.3). Также как и вся речная зона в целом, макробиотопы в ее
пределах характеризуются стабильными комплексами экологических параметров (структура
потока, глубины и скорости течения, рельеф дна, температурный режим и т.д.). Экологическая
стабильность достигается за счет формирования и функционирования русловых форм и их
населения в течение длительного периода времени.
Таблица 4.3. Макробиотопы малых водотоков Западной Камчатки (определения по [Чеботарев,
1964; Михайлов, 1977] с добавлениями)

Более низким, по сравнению с образованием русловых форм, уровнем проявления
русловых процессов является образование мозаики русловых единиц, или биотопов.
Разнообразие биотопов по большей части обусловлено грядовым движением наносов либо
неровностями коренного ложа внутри русловых форм, поэтому конфигурация биотопов может
изменяться после каждого паводка. При этом биотопы могут формироваться в условиях непре-
рывного воздействия поверхностного речного (ручьевого) стока на дно, когда воздействие
потока не прекращается в межень (центральные биотопы), либо в условиях периодического
воздействия потока на обсыхающих в межень боковых частях русла (парафлювиальная часть
речной долины по [Stanford et al., 2005]). В последнем случае формируются периферические
биотопы, в межень подпитывающиеся от стока со склонов долин, подземных вод, или во время
паводков.
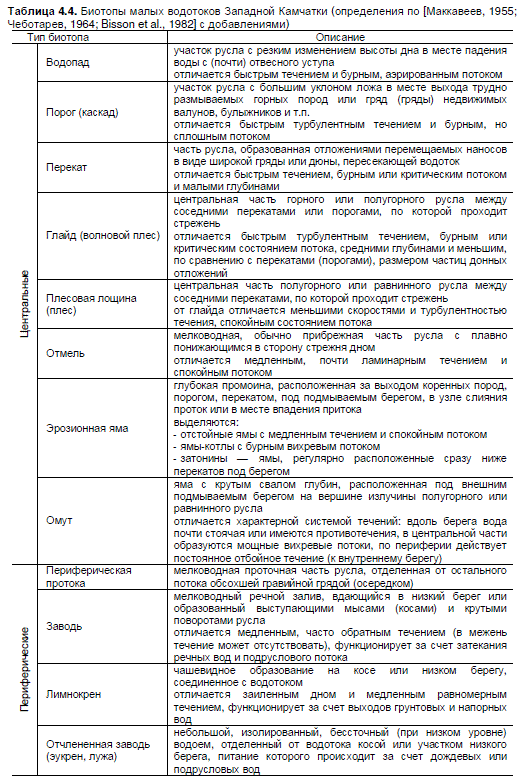
Типизация биотопов (табл. 4.4) строится на основе различий участков русел по стан-
дартным гидролого-морфологическим параметрам — прежде всего глубине и скорости течения
(Bisson et al., 1982); также учитываются продольный и поперечный микроуклон русла, крупность
частиц грунта, состояние потока и турбулентность потока (Тиунова, 2008; Stream inventory…,
1996; Church et al., 2001; Marcus et al., 2003; Wantzer et al., 2005; Moir, Pasternack, 2008).
Состояние потока может изменяться на небольшом отрезке от спокойного (плавные формы
водной поверхности) до критического (неровные формы водной поверхности) или бурного
(стоячие волны). Количественно оно выражается через число Фруда, значение которого зависит
от глубины и скорости течения (Михайлов и др., 2005). Турбулентность потока определяет ин-
тенсивность беспорядочных вихревых колебаний воды (отсутствие колебаний означает
ламинарность потока), количественно она выражается через число Рейнольдса, значение кото-
рого зависит от вектора скорости и динамической вязкости воды (Михайлов и др., 2005).
Применение комплекса параметров четко разграничивает русло на мозаику гидролого-
геоморфологических единиц.
При необходимости, деление среды на элементы можно продолжить до уровня микро-
биотопов. Например, перекат равнинной реки можно разделить на слив (или охвостье плеса —
начало косы переката), мелководный гребень (седловину), корыто (наиболее глубокую часть
гребня, через которую проходит стрежень), верхние и нижние смоченные гравийные побочни и
подвалье, т.е. обращенный вниз по течению скат гребня (Маккавеев, 1955). Прибрежная отмель
равнинного русла разделяется на мелководье и пограничную зону с направленным течением и
свалом глубин (Павлов и др., 2007). Однако в реальных условиях неоднородность свойств
потока и рельефа речного дна на микроуровне проявления не поддается точной
пространственной оценке, т.к. расположение отдельных камней, гряд или промоин в руслах
может быть случайным.
43
Таблица 4.4. Биотопы малых водотоков Западной Камчатки (определения по [Маккавеев, 1955;
Чеботарев, 1964; Bisson et al., 1982] с добавлениями)
Важность микроформ рельефа в распределении рыб в наибольшей степени проявля-
ется в неблагоприятных условиях, например, при критических скоростях течения. Более полное
освоение пространства и кормовой базы в пределах макробиотопа достигается за счет рассе-
ления гидробионтов по обособленным микробиотопам - укрытиям, т.е. локальным
местообитаниям отдельных особей, где происходит замедление потока вследствие его
столкновения с каким-то препятствием (Hunter, 1973). В качестве препятствия могут высту-
пать отдельные крупные камни или их гряды, нерегулярно расположенные по реке затопленные
бревна, древесные заломы, сложные завалы-дамбы, плотные куртины водных растений, сви-
сающие в воду ветки и корни околоводных растений, оторванные от берега куски дерна и т.п.
Вокруг препятствия задерживается часть влекомых по дну наносов, что приводит к возникнове-
нию дополнительных гряд мелкого субстрата и подпруживанию потока. В таких местах
создаются вихревые течения, которые используются рыбами для экономии энергии.
Разнообразие возможных для использования типов укрытий (микробиотопов) определя-
ется стереотипностью поведения организмов (Веселов, 1993). Использование рыбами
локальных областей вихревых течений в быстром ламинарном потоке подтверждают лабора-
торные опыты в аквариальных установках (Masahiko et al., 1997). Дополнительным стимулом
обитания молоди рыб в укрытиях является задержка в водоворотных течениях сносимых пото-
ком кормовых организмов (дрифт беспозвоночных) (Bradford, Higgins, 2001) и возможность
ускользнуть от хищника в непредсказуемую сторону вместе с одной из вихревых струй (Giger,
1973).
Для молоди лососей на разных стадиях жизненного цикла могут быть характерны некон-
тактные или контактные позиции в укрытиях. В первом случае рыбы лавируют в водяном
подпоре от препятствия (или за ним), оставаясь в "точке" гидродинамического равновесия,
постоянно подрабатывая плавниками; во втором случае рыба упирается плавниками и (или)
брюхом в субстрат (Веселов, 1993; Живоглядов, 2004; Hurtman, 1985).
Подходящие укрытия часто формируются на первоначально слабозаселенных молодью
рыб глайдах и перекатах горных русел. Утверждается (Hooper, 1973; Wesche, 1985), что нали-
чие большого количества обособленных укрытий достоверно повышает рыбопродуктивность
малых рек в десятки раз. О важности укрытий для молоди лососевых рыб на горных и полугор-
ных руслах говорит еще и тот факт, что ликвидация более 80 % удобных микробиотопов при
мелиоративных работах приводит к сокращению плотности обитания более чем в 3 раза
(Леман и др., 2000; Elser, 1968; Swales, 1982). Помимо собственно водных укрытий лососевые
также используют затененные участки под надводными выступами берегов и прибрежной рас-
тительностью. Многими исследователями (Baldes, Vincent, 1969; Giger, 1973; Gabe, Francis,
1998 и др.) показано, что "надводные укрытия" необходимы для молоди многих видов лососей.
Смотрите также






