Морфометрические различия
Поскольку кижуч откладывает икру в места выхода грунтовых вод, имеющих стабильный
температурный и гидрохимический режим (Грибанов, 1948; Леман, 2003), эмбриональное раз-
витие вида во всех зонах происходит в идентичных условиях. Именно с этим связано
отсутствие различий по меристическим признакам, число которых зависит от температуры во
время эмбриогенеза (Павлов, 1989). Из 9 исследованных счетных параметров (ll, D, A, P, V, rb.,
sb., pc, vert.) достоверные различия между выборками из разных частей бассейна наблюдаются
только по количеству жаберных тычинок; при этом величина различий минимальная и, видимо,
не имеет экологической обусловленности. Среднее число жаберных тычинок у молоди с раз-
ветвленных горных русел составляет 21,6, изменяясь от 20 до 23, с равнинных русел — 22,0 (от
20 до 24), с "болотных" русел — 21,8 (от 20 до 23), с устьевого участка — 21,7 (от 19 до 23).
Диапазон колебания величин всех признаков находится в пределах изменчивости, характерной
для вида (Черешнев и др., 2002).
115
Таблица 7.5. Показатели линейного роста молоди кижуча из бассейна р. Начилова в мае-июле
2006 г, восстановленные с помощью уравнения аллометрического роста.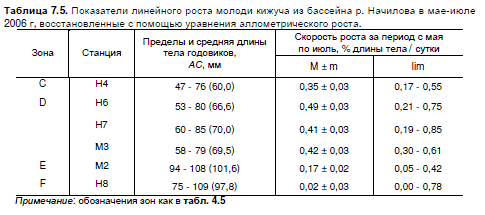
Для сравнения двухлеток кижуча из разных зон бассейна по пропорциям тела из уловов
со всех станций за 2004 г. отобрали одноразмерные выборки, достоверно не различавшиеся по
распределениям длины тела. Это позволило исключить влияние аллометрического роста на
результаты сравнения. Диапазоны изменчивости и средние значения пластических признаков
одноразмерной молоди из разных зон приведены в таблице 7.6.
Двухлетки кижуча отличаются оседлым образом жизни (Murphy et al., 1986; Quinn,
Peterson, 1996), вследствие чего на формирование их внешнего облика (пластических
признаков) на протяжении длительного времени оказывают воздействие местные специфиче-
ские условия среды обитания. Попарное сравнение морфометрических индексов двухлеток
кижуча (распределение индексов было изначально проверено на соответствие нормальному)
показало наличие достоверных различий между выборками из разных зон по 9—19 из 25
возможных сочетаний сравниваемых пар (табл. 7.7). Объединение выборок в одну привело к
потере симметрии и одновершинности распределения 17 индексов и лишь длина головы (с),
длины челюстей (lm, lmx, lmd), высоты тела (H, h), длина анального плавника (lA) и
антевентральное расстояние (аV) не утратили нормальности распределения.
Размах изменчивости признаков, выражающийся через отношение достоверно отличаю-
щихся пар сравнения к числу сравниваемых пар, у зонально подразделенных выборок невысок.
Это может быть объяснено генетическим единством нерестового стада кижуча малой реки. Тем
не менее, наличие достоверных различий по большинству параметров свидетельствует о мор-
фологической дифференциации одноразмерной молоди из разных зон бассейна. Очевидно, что
морфологические особенности молоди кижуча из летних пространственных группировок отра-
жают в своей основе адаптации представителей этих группировок к факторам среды обитания
(скорости течения, турбулентности потока и т.п.). Наибольшие различия выборок проявляются в
высоте спинного (hD) и длине грудных плавников (lP), положении анального (aA) и парных
плавников (P-V, V-A), а также длине рыла (ao); наименьшие — в высоте тела (H, h), положении
спинного плавника (aD) и пропорциях элементов головы (длина верхней челюсти, высота глаза,
длина жаберной крышки).
При многофакторном разделении двухлеток кижуча, относящихся к группировкам разных
зон (численность использованных в анализе выборок приведена в таблице 7.6), в функцию
многомерной дискриминации включаются 18 из 25 индексов морфометрических промеров. Рас-
пределение всех индексов соответствует нормальному, асимметрия и эксцесс незначимы.
Дисперсия 8 из 18 дискриминирующих параметров (по статистике Лямбды Уилкса) достоверно
различается во всех группах сравнения. К наиболее значимым разделяющим индексам отно-
сятся признаки, которые проявляли максимальную изменчивость при попарном сравнении:
длина грудных (lP), высота спинного (hD) и анального (hA) плавников, расстояние между пар-
ными плавниками (P-V), положение анального плавника (aA), длина и ширина рыла (ao и io).
Максимальное значение Лямбды Уилкса, величина которой демонстрирует степень общности
по параметру, составляет 0,021. Объекты (отдельные особи) включаются в состав групп, выде-
ленных по зональному принципу, с вероятностью 97 % (0,05≤ р ≤0,01).
116
Таблица 7.6. Морфометрическая характеристика кижуча 1+, выловленного в разных зонах бассейна р. Начилова в июле 2004 г. (индексы в % от
длины тела)
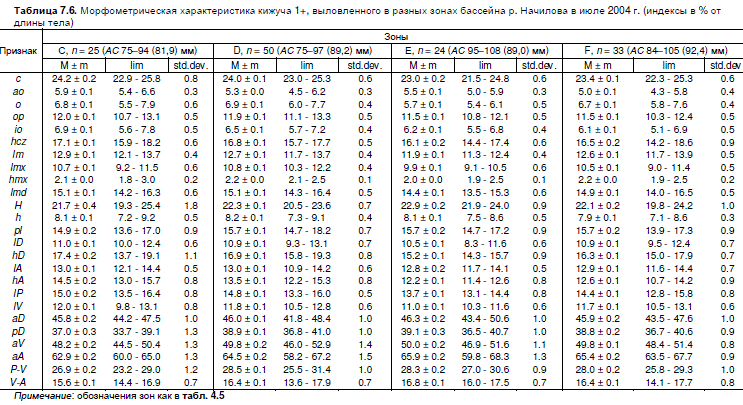
Таблица 7.7. Оценки достоверности различий между морфотипами кижуча 1+, выловленного в
июле 2004 г. в разных зонах бассейна р. Начилова, по критерию Стьюдента
Пары сравниваемых выборок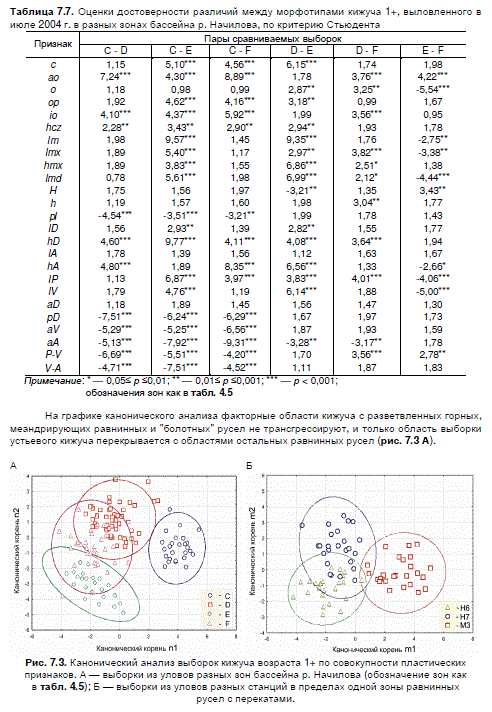
обозначения зон как в табл. 4.5
На графике канонического анализа факторные области кижуча с разветвленных горных,
меандрирующих равнинных и "болотных" русел не трансгрессируют, и только область выборки
устьевого кижуча перекрывается с областями остальных равнинных русел (рис. 7.3 А).
А Б
Рис. 7.3. Канонический анализ выборок кижуча возраста 1+ по совокупности пластических
признаков. А — выборки из уловов разных зон бассейна р. Начилова (обозначение зон как
в табл. 4.5); Б — выборки из уловов разных станций в пределах одной зоны равнинных
русел с перекатами.
118
Обоснованность выделения летних пространственных группировок двухлеток кижуча
подтверждается результатами сравнения выборок, выловленных на разных станциях в преде-
лах одной зоны меандрирующих равнинных русел с перекатами. Анализ морфометрии кижуча
длиной 78—102 мм из июльских уловов станций Н6, Н7 и М3 (выборки по 24—27 экз.) за 2005 г.
показал, что выборки достоверно различались по 5—13 из 25 сравниваемых пар, однако все
различия проявлялись лишь в какой-то одной паре сравнения. Многофакторное разделение
выборок было возможно лишь на основании длины хвостового стебля (pl), дисперсии прочих
параметров перекрывались. Объекты включаются в состав выделенных групп недостоверно,
р ≤ 0,76 (рис. 7.3 Б).
Межгодовые изменения внешних признаков двухлеток кижуча внутри зоны меандри-
рующих равнинных русел также меньше различий между пестрятками из разных зон. В уловах
станции М3 изменения пластических признаков в выборках за 2003, 2004 и 2005 гг. (по 25 экз.)
затронули 18 из 25 параметров. Дисперсия 10 индексов (lmx, aV, lm, h, io, lmd, H, hD, pl и lР)
различалась во всех группах сравнения, однако она была в 2 раза уже, чем при анализе
выборок из разных зон — максимальная Лямбда Уилкса составила 0,049 против 0,021
соответственно. Объекты включаются в состав выделенных групп недостоверно, р ≤ 0,85.
Смотрите также






