СЫРЬЕВЫЕ РЕСУРСЫ МОРСКИХ РЫБ САХАЛИНА И КУРИЛЬСКИХ ОСТРОВОВ: СОСТАВ, СОВРЕМЕННОЕ СОСТОЯНИЕ ЗАПАСОВ, ИХ МНОГОЛЕТНЯЯ ИЗМЕНЧИВОСТЬ
Сахалин и Курильские острова (вместе с японским о. Хоккайдо) представляют собой уникальную систему морских островов. Протяженность этой островной дуги, точнее только ее российской части, составляет около 2200 км, что сопоставимо с максимальной протяженностью Охотского моря с юго-запада на северо-восток - до 2500 км. В биогеографическом смысле значительная часть морских и океанических акваторий этой островной системы относится к Японской низкобореальной подобласти. Лишь воды северной части Сахалина и северных Курильских островов входят в охотоморскую часть Алеутско-Камчатской высо-кобореальной провинции (Парин, 1968; Несис, 1982; Шунтов, 1985). В целом рассматриваемая группа островов отделяет друг от друга 3 крупномасштабных водоема: Японское и Охотское моря и северо-западную часть Тихого океана.
Специфичность географического расположения в совокупности со значительной суммарной протяженностью данной островной системы предопределили в ее отдельных районах существенные различия в климате, океанологическом режиме, а также наложили отпечаток на формирование разнообразных биотопов для морских гидробионтов. В состав крупных морских биотопов, образовавшихся вблизи рассматриваемой группы островов, входят, например, прибрежные, мелководные зоны, шельф, эпипелагиаль, а также мезопелагиаль и непосредственно материковый склон. Это, в свою очередь, отразилось на формировании в исследуемой акватории очень богатой по видовому обилию ихтиофауны, в том числе и на значительном для дальневосточных морей России разнообразии промысловых видов рыб. В частности, в настоящее время в Сахалино-Курильском регионе число видов рыб, по которым разрабатываются прогнозы ОДУ, достигает 30 (без учета видового состава камбал, бычков, скатов), тогда как в зоне Приморья - только 10, в северной части Охотского моря - 13, у западной Камчатки - 15, в западной части Берингова моря - 19. Уместно напомнить, что в самом начале 20-го столетия у берегов Сахалина, например, были разведаны и осваивались промыслом запасы всего 6 видов рыб: сельди, трески, палтуса, кеты, горбуши и осетра (Шмидт, 1905). Более менее в окончательном виде сырьевые ресурсы морских рыб Сахалино-Курильского региона были выявлены лишь к концу 20-го века. В целом история отечественных рыбохозяйственных
исследований в водах, прилегающих к Сахалину и Курильским островам, охватывает длительный, более чем вековой, период. Начало этим научным исследованиям было положено П.Ю.Шмидтом (1905). В дальнейшем на протяжении всего столетия, в особенности со 2-й половины 40-х гг., периодически стали публиковаться научные труды, в которых обобщалась комплексная информация о рыбных ресурсах рассматриваемого района, имевшаяся на соответствующий отрезок времени. Как правило, они касались характеристики сырьевых ресурсов морских рыб либо отдельных районов Сахалина и Курильских островов, либо какой-то группы видов или рыбных сообществ (Таранец, 1937; Черфас, 1946; Борисов, 1949; Веденский, 1949; Линдберг, 1954, 1959; Моисеев, 1954; Пробатов, Лестев, 1954; Расс, 1955, 1956, 1959; Сафронов, Никифоров, 1981; Борец, 1985, 1990; Гаврилов и др., 1988; Дудник, Долганов, 1992; Дудник и др., 1995; Зверькова, Пушникова, 1996; Радченко и др., 1997; Иванов, 1998; Ильинский, 1998; Гаврилов, 1998; Каредин, 1998; Шунтов и др., 1998а, б; Великанов, 1999). Кроме того, в последнюю четверть 20-го века были опубликованы ряд монографий и сборников статей, в которых так или иначе также рассматривались, комплексно и пообъектно, ресурсы морских рыб, текущее состояние запасов и их многолетняя изменчивость в водах Сахалина и Курильских островов (Новиков, 1974; Фадеев, 1984, 1987; Шунтов, 1985; Биологические ресурсы Тихого океана, 1986; Гидрометеорология и гидрохимия морей, 1993; Промысловые рыбы ..., 1993; Борец, 1997; Промыслово-биологические исследования рыб ..., 2000).
Из этого краткого обзора можно заключить, что к концу 20-го столетия в основном завершился разведочный этап исследований сырьевых ресурсов рыб в водах Сахалина и Курильских островов. При этом был установлен видовой состав промысловых и потенциально промысловых рыб основных экологических групп: пелагических, донных, глубоководных, в том числе мезопелагических. Были изучены основные черты биологии массовых видов, для многих промысловых популяций прослежена многолетняя динамика численности и ее флюктуации, накоплены статистические ряды ежегодных уловов, определены промысловые возможности большинства популяций (или единиц запаса). В связи с развитием мер промыслового регулирования в последнюю четверть века для подавляющего большинства контролируемых мониторинговыми исследованиями единиц запаса установлены более менее достоверные величины допустимого улова. Таким образом, накопленная к настоящему времени значительная по объему и содержательности информация о сырьевых ресурсах морских рыб Сахалина и Курильских островов позволяет осуществить определенные обобщения и выявить некоторые особенности, присущие промысловой ихтиофауне рассматриваемого региона.
Цель настоящей работы заключается в современной характеристике сырьевых ресурсов морских рыб Сахалина и Курильских островов с кратким обобщением накопленных материалов по видовому составу, состоянию запасов и их многолетней изменчивости.
Информационной основой статьи послужили несколько источников. Во-первых, использованы результаты донных траловых съемок и постановок донных жаберных сетей, выполненных СахНИРО в рамках исследовательских программ на шельфе и материковом склоне Сахалина и Курильских островов в 80-90-е гг. Кроме того, широко использовалась разнообразная опубликованная информация. В частности, для экспертной оценки изменений видового состава доминирующих видов рыб по отдельным районам за последние 50 лет помимо вышеупомянутых
публикаций использовались и другие (Пробатов, Шелегова, 1968; Кани-дьев, 1972; Золотов, 1986; Fishes and Marine invertebrates of Hokkaido ..., 1991; Шунтов и др., 1993; Тарасюк, 1994; Лапко, 1996; Савин, 1998; Зверь-кова, 1999; Худя, 1999; Zhigalin, Belayev, 1999). Также использованы официальные данные о величинах ОДУ различных видов рыб для отдельных промысловых зон и подзон Сахалино-Курильского региона в 19912001 гг., основанные на материалах сотрудников СахНИРО и ТИНРО-центра. Данные по статистике вылова рыб взяты из материалов Сах-НИРО, ТИНРО-центра и Сахрыбвода.
В структуре морских биологических ресурсов дальневосточной ИЭЗ России сырьевые запасы рыб занимают соответствующее представителям верхних трофических уровней ведущее место, составляя по оценкам последних лет почти 81 % суммарной величины ОДУ (рис. 1). Примерно такую же величину имеют рыбы в общем объеме мирового вылова, которая колеблется, по данным П.А.Моисеева (1969), в пределах 81-86 %. В Сахалино-Курильском регионе сырьевые ресурсы морских рыб также занимают лидирующее положение по сравнению с другими категориями водных биоресурсов, составляя в 2000-2001 гг. почти 50 % суммарного ОДУ (рис. 2). Существенны здесь также ресурсы кальмаров и водорос-лей-макрофитов, совместная доля которых - более 40 %. Значительна роль морских рыб и в промысловых уловах водных биоресурсов Сахалина и Курильских островов в современный период. В годовом улове их доля изменяется от 39,1 % (1999 г.) до 53,5 % (2000 г.), эти вариации обусловлены изменением уловов лососей (прежде всего горбуши) в четные и нечетные годы (рис. 3).
Рис. 1. Соотношение рыб, беспозвоночных и водорослей в структуре морских биоресурсов в дальневосточной ИЭЗ России в 20002001 гг.
Fig. 1. Proportion of fish, invertebrates and algae in marine biore-sources structure in the Far East exclusive economic area of Russia in 2000-2001
Phc. 2. CooTHomeHHe pu6, 6ec-n03B0H0HHbix h BOAopoc^eô b CTpyK-Type MopcKHx 6HopecypcoB b caxa-^HHo-KypH^bCKoM pafioHe b 20002001 rr.
Fig. 2. Proportion of fish, invertebrates and algae in marine biore-sources structure in Sakhalin-Kurile region in 2000-2001
Морские экосистемы Сахалина и Курильских островов характеризуются рядом особенностей, которые оказали значительное влияние на формирование качественного состава промысловой ихтиофауны рассматриваемого района. Прежде всего это сравнительно узкий островной шельф. Как известно, наличие хорошо развитого шельфа играет чрезвычайно важную роль в формировании его биологических ресурсов (Моисеев, 1969). В то же время в Охотском море, например, восточносахалинский и южноохотоморский шельфы (о. Хоккайдо и Курильские острова) суммарно составляют всего 18 % общей площади шельфовой зоны этого водоема (Благодеров, Маркина, 1986). Вследствие узости островного шельфа зона больших глубин (материковый склон) расположена здесь весьма близко от берега. Кроме того, прилегающие к островам морские акватории в основном расположены в южной части дальневосточной экономической зоны и находятся под активным влиянием теплых течений системы Куросио. По мнению В.И.Радченко с соавторами (1997), южная часть Охотского моря и тихоокеанские воды Курильских островов - один из наиболее динамичных районов экономической зоны России. В эпипелагиали Сахалино-Курильского региона трансформированные воды относительно изолированного от океана Охотского моря взаимодействуют с водами, приносимыми течением Соя из Японского моря и Курило-Камчатским течением - из Берингова. К важным океанологическим особенностям этого района относятся такие как присутствие на акваториях прикурильских районов субарктических и субтропических вод, мозаичность распределения гидрологических характеристик, наличие большого числа протяженных фронтальных зон. Последнее обусловливает образование плотных концентраций планктонных и нектонных организмов, в том числе пелагических рыб и кальмаров, служащих объектами промысла (Новиков, 1986).
Рис. 3. Соотношение уловов рыб, беспозвоночных и водорослей в сахалино-куриль-ском районе в 1999 г. (А), 2000 (Б) и 19992000 гг. (В)
Fig. 3. Proportion of fish, invertebrates and algae catches in Sakhalin-Kurile region in 1999 (A), 2000 (Б) and 1999-2000 (В)
Кроме того, воздействие процессов, происходящих в Сахалино-Курильском регионе, на условия среды в северо-западной Пацифике в целом позволяет говорить не столько о переходном, сколько о ключевом статусе этого района. Здесь не только взаимодействуют, но и вновь формируются потоки, во многом определяющие океанологические условия и особенности структуры и функционирования сообществ гидробионтов на близлежащих акваториях (Радченко и др., 1997).
Как показали многолетние научные наблюдения и исследования, в целом в этом регионе промысловая ихтиофауна сформирована тремя основными группами (сообществами) рыб: донными и придонными шельфовыми, пелагическими, глубоководными. Донные и придонные рыбы шельфа представлены главным образом тресковыми, камбаловыми, бычками, песчан-1125

ковыми и скорпеновыми. Пелагические - большим числом семейств (сельдевые, анчоусовые, лососевые, корюшковые, макрелещуковые, тресковые, терпуговые, скумбриевые и тунцовые). Группу рыб материкового склона и больших глубин составили представители скатовых, батилаго-вых, светящихся анчоусов, моровых, долгохвостых, бельдюговых, скорпено-вых и камбаловых.
В связи с крайне ограниченной площадью островного шельфа запасы донных рыб здесь сравнительно небольшие (менее 12 %). Разведанные запасы глубоководных рыб, осваиваемых промыслом в настоящее время, тоже относительно невелики (около 9 %). Конечно, при условии включения в состав сырьевой базы рыболовства выявленных в последнее время запасов мезопелагических рыб: серебрянки в западной части Охотского моря, лампаниктов, батилагов и диафуса теты в тихоокеанских водах Курильских островов (Ильинский, 1994, 1998; Каредин, 1998), -доля этой группы видов может существенно возрасти. Однако перспективы промыслового освоения ресурсов мезопелагических рыб пока представляются весьма отдаленными и проблематичными по целому ряду причин: техническим (технологическим), из-за недостаточной изученности объектов, недостаточного понимания роли этих видов в морских экосистемах (Кашкин, 1989; Каредин, 1998). Таким образом, наиболее многочисленны в морских водах Сахалина и Курильских островов пелагические виды рыб, доминирующие по биомассе и составляющие (без лососей) в среднем 66,7 % (рис. 4).
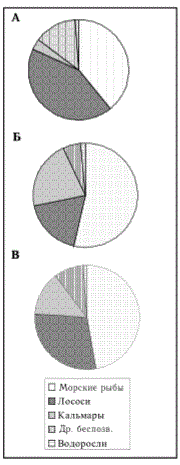
Рис. 4. Соотношение донных пелагических, анадромных и глубоководных рыб в районе северных Курильских островов (А), южных Курильских островов (Б), восточного Сахалина (В), западного Сахалина (Г), в целом (Д) в 20002001 гг.
Fig. 4. Proportion of bottom, pelagic, anadromous and deep-water fish in regions of the North Kuriles Islands (A), South Kuriles (Б), East Sakhalin (В), West Sakhalin (Г), integrally (Д) in 2000-2001
Эпипeлaгичecкoe cooбщecтвo pыб пpeдcтaвлeнo здecь мк ceвepoбo-peaльными видaми (ceльдь, минтaй, мoйвa, тиxooкeaнcкиe лococи - гopбy-ma, кeтa и дp.), тaк и пepиoдичecки мигpиpyющими cюдa ввд^ми cyбтpo-пичecкoгo кoмплeкca (дaльнeвocтoчнaя capдинa, caйpa, cкyмбpия, япoнc-кий aнчoyc), a тaкжe oбычнo oбитaющими и paзмнoжaющимиcя в тpoпи-чecкиx вoдax Tиxoгo oкeaнa двумя видaми тундов - гoлyбым и пoлoca-тым. Cлeдyeт oтмeтить, чтo cyбтpoпичecкиe виды pb^ coвepшaют cвoи мигpaции, oбpaзyя бoльшиe cкoплeния пpoмыcлoвoгo xapaктepa, œ толь-кo в тиxooкeaнcкиe вoды южныx Kypильcкиx ocтpoвoв, нo и к пoбepe-жью Caxaлинa, в Taтapcкий пpoлив (Tapaнeц, 1937; Pyмянцeв, 1947; Шyнтoв, 1967; Дapдa, 1968; Beликaнoв, Tapacюк, 1981; Фaдeeв, 1984; Дyдa-peв, Keня, 1986; Bacилeнкo, Шepшeнкoв, 1997; Beликaнoв, в пeчaти) и в бoлee xoлoдныe oxoтoмopcкиe вoды ocтpoвa, нepeдкo дocтигaя ero ceвe-po-вocтoчныx бepeгoв. Taк, пo дaнным A.Я.Tapaнцa (1937), в 30-e гг. дaльнeвocтoчнaя capдинa (ивacи) вcтpeчaлacь в yлoвax cтaвнoгo нeвoдa в нeкoтopыx зaливax ceвepo-вocтoчнoгo Caxaлинa (Hracra^ Чaйвo). В кoнцe 90-x гг. в югo-зaпaднoй чacти Oxoтcкoгo мopя шиpoкoe parapoCT-paнeниe имeл япoнcкий aнчoyc в cвязи c нaблюдaвшимcя cyщecтвeн-ным pocтoм eгo чиcлeннocти (Шyнтoв и дp., 1998a). В чacтнocти, этoт вид cтaл вcтpeчaтьcя в бoльшиx кoличecтвax нe толь^ в вoдax юго-вocтoчнoгo Caxaлинa, гдe в 50-e гг. тaкжe нaблюдaлиcь eгo cкoплeния (Дapдa, 1968), нo и нa ceвep вдoль Caxaлинa дo 53o c.ш. O6 этoм cвидe-тeльcтвyют peзyльтaты дoнныx тpaлoвыx cъeмoк, ocyщecтвлeнныx Cax-HOTO в ceнтябpe-oктябpe 1998-1999 гг. (Beликaнoв, в пeчaти).
В цeлoм жe в пocлeдниe гoды дoля пeлaгичecкиx pыб в cтpyктype pыбныx pecypcoв в oтдeльныx paйoнax (пpoмыcлoвыx зoнax и пoдзo-нax) вecьмa знaчитeльнo кoлeблeтcя, cocтaвляя oт 7,7 в Bocтoчнo-Caxa-линcкoй пoдзoнe дo 83,5 % в Южнo-Kypильcкoй зoнe (pœ. 4, a-г). Bпoлнe oчeвиднo, что выявлeнныe cooтнoшeния paзличныx экoлoгичecкиx фупп pыб (cooбщecтв) в нeкoтopыx paйoнax вpeмя oт вpeмeни будут cyrn,e-cтвeннo измeнятьcя. В чacтнocти, мoжнo пoлaгaть, нaпpимep, чтo в Boc-тoчнo-Caxaлинcкoй пoдзoнe дoля пeлaгичecкиx pыб в 80-e гг. былa, нeчнo, нecpaвнимo вышe в cвязи c выcoкoй чиcлeннocтью нa тот пepиoд минтaя и capдины (Шунтов и дp., 1993; Звepькoвa, 1999; Zhigalin, Belayev, 1999). Пpичины тaкoй вpeмeннoй нeycтoйчивocти cooтнoшeния paзлич-ныш экoлoгичecкиx гpyпп pыб кpoютcя, мк извecтнo, в ocнoвнoм в ^уп-нoмacштaбнoй измeнчивocти cиcтeмныx пpиpoдныx пpoцeccoв (Шyнтoв, 1993, 1994; Kляштopин, Cидopeнкoв, 1996; Шyнтoв и дp., 1997).
Oднoй из вaжнeйшиx ocoбeннocтeй измeнeний климaтa Ceвepнoгo пoлyшapия в пpoшeдшeм cтoлeтии cчитaютcя eгo дoлгoпepиoдныe вapи-aции. Kaк ycтaнoвлeнo, пepexoд oт фaз xoлoднoгo климaтa к тeплoмy coпpoвoждaлcя измeнeниeм пapaмeтpoв тepмoдинaмичecкoгo cocтoяния cиcтeмы, oпpeдeлeннoй пepecтpoйкoй aтмocфepнoй циpкyляции. Пpи этoм пoля пpизeмнoй тeмпepaтypы вoздyxa, aтмocфepнoгo дaвлeния нa ypoвнe мopя и тeмпepaтypы пoвepxнocти oкeaнa иcпытывaли coглacoвaнныe климaтичecкиe вoзмyщeния (Бышeв и дp., 1997). В cвязи c этим вaжнoe знaчeниe пpидaeтcя динaмикe и пpeoблaдaнию нa paзныx этaпax oпpeдe-лeнныx фopм aтмocфepнoй циpкyляции, влияющиx в cвoю oчepeдь нa ocoбeннocти циpкyляции вoд, т^. глaвныx cocтaвляющиx климaтooкea-нoлoгичecкиx эпox. Bce этo oтpaжaeтcя нa фopмиpoвaнии лaндшaфтнoгo и биoтoпичecкoгo oкpyжeния гидpoбиoнтoв и чepeз нeгo пpeлoмляeтcя в динaмикe чиcлeннocти пoпyляций и cтpyктypы cooбщecтв. В чacтнo-cти, в нaчaлe 90-x гг. в дaльнeвocтoчныx мopяx Poccии нaчaлиcь ^уп-
1982 <и са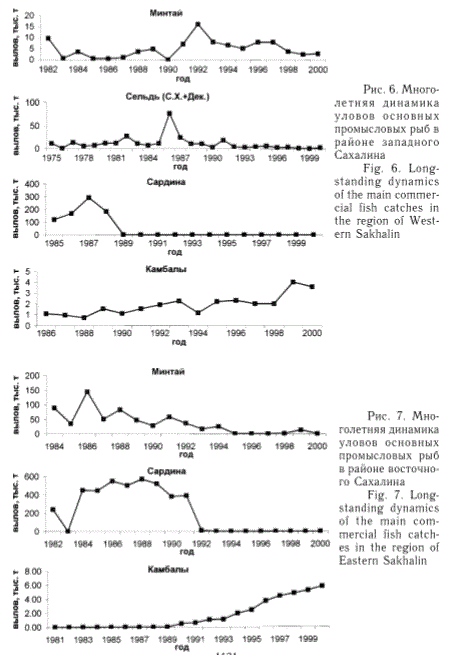
ные экосистемные перестройки, вызванные глобальными изменениями климатоокеанологических и космо-геофизических факторов. Эти естественные процессы охватили прежде всего и в основном динамичные пелагические сообщества и ярко проявились во флюк-туациях численности таких массовых рыб, как минтай, дальневосточная сардина, сельдь и др. (Шунтов и др., 1997).
Предпринимаемые усилия в исследовании данной проблемы уже выявили немало примеров влияния долгопериодной изменчивости атмосферной циркуляции над Дальневосточным регионом на термический режим и динамику морских вод в разных районах, в том числе на акваториях, прилегающих к Сахалину и Курильским островам (Матвеев, Мороз, 1997; Павлы-чев, 1997; Хен, 1997; Шатилина, 1997, 1998; Шунтов и др., 1997; Самко и др., 1998). Согласно Т.А.Шатили-ной (1998), общая схема взаимодействия этих процессов может выглядеть следующим образом. В летний период усиление течения Ой-ясио хорошо согласуется с процессами, протекающими над Охотским морем, в том числе с интенсивностью формирования холодной ложбины над восточной частью Охотского моря. Эти атмосферные процессы "отвечают" за аномальность термики и циркуляции вод Японского, Охотского морей и прилегающей части Тихого океана, так как они регулируют поступление тепла на север с ветвями Куросио.
Значительная долгопериодная изменчивость океанологических условий, присущая вышеперечисленным морским водоемам, и в частности акватории, окружающей Сахалин, может быть проиллюстрирована данными рис. 5, любезно предоставленного заведующим океанологической лабораторией СахНИ-РО Г.А.Кантаковым. Как видно на рис. 5, на известном стандартном
Aнтoнoвcкoм paзpeзe у югo-зaпaднoгo пoбepeжья Caxaлинa зa пoчти 80 лeт нaблюдeний oтмeчeнo нecкoлькo пepиoдoв бoлee xoлoднoгo и бoлee тeплoгo тepмичecкoгo peжимa вoд. Haпpимep, в пpoшлoм cтoлeтии пoни-жeнный тeмпepaтypный фoн вoд нaблюдaлcя в 20, 50 и 80-e гг., a пoвышeн-ный - в 30, 60 и 90-e гг.
Измeнчивocть климaтooкeaнoлoгичecкиx ycлoвий, в тoм чиcлe тeм-пepaтypнoгo peжимa, кaк yжe oтмeчaлocь, oкaзывaeт cyщecтвeннoe влия-нте нa биoлoгичecкyю пpoдyктивнocть вoд, что oтpaжaeтcя нa ycпeшнocти вocпpoизвoдcтвa pыб и измeнeнии ypoвня иx зaпacoв. В cвязи c этим yмecтнo oтмeтить, чтo paнee были ycтaнoвлeны дocтoвepныe кoppeляци-oнныe cвязи чиcлeннocти пoкoлeний pыб c paзличными гидpoмeтeopoлo-гичecкими фaктopaми для пoпyляций минтaя, нaвaги, мoйвы, дaльнeвocтoч-нoй пecчaнки, жeлтoпepoй кaмбaлы, paзмнoжaющиxcя у бepeгoв Caxaлинa (Звepькoвa, 1973; Beликaнoв, 1986, 1991; Caфpoнoв, 1986; Худя, 1990; Tapa-cюк, 1994), a тaкжe - для caйpы, cкyмбpии, capдины, япoнcкoгo aнчoyca, paзмнoжeниe кoтopыx пpoиcxoдит в вoдax тeплoгo тeчeния Kypoœo (Co-кoлoвcкий, 1972; Бeляeв, 1986; Швимв, 1986; Caблин, Ивaнoв, 1986).
В cвязи c вышeизлoжeнным впoлнe зaкoнoмepнo, чтo cocтaв и личecтвo дoминиpyющиx видoв pыб в paйoнe Caxaлинa и южныx Ky-pильcкиx ocтpoвoв пepиoдичecки cyщecтвeннo измeняютcя (тaбл. 1). Taк, в Taтapcкoм пpoливe чиcлo дoминиpyющиx видoв pыб измeнялocь c 50-x пo 90-e гг. 20-го cтoлeтия c 2 дo 4, у бepeгoв вocтoчнoгo Caxaлинa -oт 1 дo 5, у южныx Kypильcкиx ocтpoвoв - c 4 дo 8. У ceвepныx Kypиль-cкиx ocтpoвoв чиcлo дoминиpyющиx видoв pыб бoлee cтaбильнo, oбыч-нo пpeдcтaвлeнo 2-3 ввд^ми. Ecли гoвopить o кaчecтвeннoм cocтaвe pыб, тo oн тaкжe пoдвepжeн cyщecтвeннoй измeнчивocти. Taк, в Taтapc-кoм пpoливe в 50-e гг. дoминиpoвaли ceльдь, минтaй, cкyмбpия и aнчoyc, в 80-e - capдинa, минтaй, ceльдь и мoйвa, в 90-e - aнчoyc, тpecкa и минтaй. У вocтoчныx бepeгoв Caxaлинa в 50-e гг. пpeoблaдaли ceльдь, минтaй, пecчaнкa, aнчoyc и кaмбaлы, в 80-e - capдинa ивacи, минтaй, œc-чaнкa и гopбyшa, в 90-e гг. - гopбyшa, южный oднoпepый тepпyг, aнчoyc и кaмбaлы. У южныx Kypильcкиx ocтpoвoв в 50-e гг. дoминиpoвaли тaкиe виды, кaк гopбyшa, caйpa, aнчoyc и тунцы, в 70-e - в ocнoвнoм минтaй, capдинa, caйpa, cкyмбpия, южный oднoпepый тepпyг, гopбyшa, в 90-e гг. - caйpa, япoнcкий aнчoyc, тунцы, южный тepпyг, минтaй, гopбyшa. В дaннoм paйoнe нaибoльшeй дoлгoпepиoднoй ycтoйчивocтью зaпacoв xapaктepизyютcя caйpa и гopбyшa, что yжe нeoднoкpaтнo oтмeчaлocь paнee (Ивaнкoв, 1984; Hoвикoв, 1986). У ceвepныx Kypильcкиx ocтpoвoв из-мeнчивocть cocтaвa дoминиpyющиx видoв выpaжeнa в мeньшeй ^eM-ни, a caмыми мнoгoчиcлeнными видaми pыб пoчти зa вecь пepиoд нaблю-дeний были минтaй, ceвepный oднoпepый тepпyг и мaкpypycы.
Taким oбpaзoм, пepиoдичecкaя измeнчивocть cocтaвa дoминиpyю-щиx пo биoмacce pыб являeтcя xapaктepнoй ocoбeннocтью фyнкциoни-poвaния и пpoдyциpoвaния мopcкиx экocиcтeм Caxaлинa и южныx Ky-pильcкиx ocтpoвoв. Cдeлaнный вывoд мoжeт имeть вaжныe пpaктичec-киe cлeдcтвия, тaк мк этo пoлeзнo yчитывaть пpи пpoгнoзиpoвaнии œ-pьeвoй бaзы нa paзличнyю пepcпeктивy, пpи плaниpoвaнии paзвития pы-бoлoвcтвa в дaннoм peгиoнe, a тaкжe пpи плaниpoвaнии и opгaнизaции мoнитopингoвыx и пepcпeктивныx pыбoxoзяйcтвeнныx иccлeдoвaний, пpeждe вceгo мopcкиx экcпeдициoнныx, нa aквaтopияx paccмaтpивaeмoгo peгиoнa.
Bo мнoгиx cлyчaяx измeнчивocть вeличины pыбныx зaпacoв oтpa-жaeтcя нa вeличинe и cocтaвe пpoмыcлoвыx yлoвoв, xoтя, кoнeчнo, нe
та
I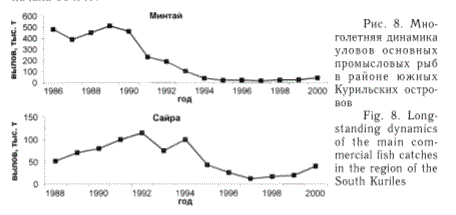
вceгдa yлoвы aдeквaт-нo пoкaзывaют cocтoя-нте зaпacoв. Haпpимep, у зaпaднoгo Caxaлинa в пocлeдниe 15-25 лeт нaибoльшиe yлoвы были пoлyчeны пo cap-динe и ceльди (pиc. 6). Пpoмыceл capдины зa-вepшeн в нaчaлe 90-x гг. в cвязи c пpeкpaщe-ниeм ee пoдxoдoв в нaшy зoнy. Улoвы ^ль-ди в ocнoвнoм были cpaвнитeльнo нeбoль-шими, и лишь в 1986 г. пpoизoшeл cкaчoк в вылoвe этoй pыбы зa cчeт oблoвa ypoжaйнo-гo пoкoлeния 1983 гoдa poждeния (Пyшникoвa, 1994). В мeньшeй мepe вылaвливaлcя минтaй, гoдoвыe yлoвы ^TOpo-гo в этот пepиoд oбыч-нo нe дocтигaли дaжe пpoгнoзиpyeмыx вeли-чин OДУ из-зa cлaбoй интeнcивнocти лoвa. В 1999 и 2000 гг. cyщe-cтвeннo yвeличилcя вылoв кaмбaл в дaн-нoм paйoнe, чтo cвязa-нo, пo-видимoмy, c швы-шeниeм cпpoca нa эту pыбy и вoзoбнoвлeни-eм aктивнoгo кoммep-чecкoгo лoвa.
У вocтoчнoгo Caxa-линa нaибoльшими yлo-вaми в мнoгoлeтнeм плaнe тaкжe xaparcrep^ зoвaлиcь capдинa ивacи (бoлee 500 тыc. т) и минтaй (дo 150 тыс т) (pиc. 7). Дoбычa этиx oбъeктoв ocyщecтвля-лacь здecь э^мдици-oнным лoвoм c пpивлe-чeниeм бoльшoгo тли-чecтвa пpoмыcлoвыx и пepepaбaтывaющиx cy-дoв. Oднaкo в нacтoя-
щее время в связи со значительным сокращением запасов былое промысловое значение этих видов в данном районе полностью утрачено. Лов сардины завершился в начале 90-х гг. из-за прекращения подходов рыбы. Величина вылова минтая с середины 90-х гг. сократилась на 1-2 порядка и в конце 90-х не превышала 13 тыс. т, сохраняя лишь локальное значение в рыболовстве. Вместе с тем после снятия запрета в 90-е гг. наблюдался рост вылова камбал, в основном за счет восстановившихся запасов популяции желтоперой камбалы зал. Терпения.
Рис. 6. Многолетняя динамика уловов основных промысловых рыб в районе западного Сахалина
Fig. 6. Longstanding dynamics of the main commercial fish catches in the region of Western Sakhalin
Рис. 7. Многолетняя динамика уловов основных промысловых рыб в районе восточного Сахалина
Fig. 7. Longstanding dynamics of the main commercial fish catches in the region of Eastern Sakhalin
Возле южных Курильских островов (рис. 8) ежегодные уловы минтая и сайры в последние 15 лет также характеризуются существенным снижением, особенно значительным по минтаю, практически на один порядок, по сайре - в основном в 2-3 раза. Однако если у минтая это обусловлено действительно сокращением запасов (Зверькова, 1999), то снижение отечественных уловов сайры связано прежде всего с причинами экономического и организационного характера. Наметившаяся тенденция некоторого увеличения уловов сайры во второй половине 90-х гг. позволяет надеяться, что со временем российский промысел этой ценной рыбы возобновится в полном объеме, а ее уловы достигнут уровня начала 90-х гг.
Рис. 8. Многолетняя динамика уловов основных промысловых рыб в районе южных Курильских островов
Fig. 8. Longstanding dynamics of the main commercial fish catches in the region of the South Kuriles
У северных Курильских островов (рис. 9) промысловые уловы минтая и трески в последние 10-15 лет также характеризуются заметным сокращением, что обусловлено именно снижением запасов данных видов рыб. Некоторое уменьшение уловов в 90-е гг. прослеживается и у камбал этого района, основу вылова которых составляет двухлинейная камбала. Однако в данном случае это связано с недоиспользованием запасов, поскольку в последнее десятилетие ни разу годовой вылов этой рыбы не достигал рекомендованной величины ОДУ. Зато вылов северного одноперого терпуга существенно возрос во вторую половину 90-х гг. в результате роста численности этой рыбы.
Для более полной характеристики современного состояния запасов морских рыб Сахалина и Курильских островов коротко приведем результаты анализа тенденций изменения запасов и прогнозируемых допустимых уловов основных промысловых видов в предыдущее десятилетие по отдельным районам, которые показали следующее.
Возле западного Сахалина (рис. 10) в 90-е гг. предыдущего столетия запасы минтая, сельди, мойвы характеризовались тенденцией снижения. Об этом свидетельствовали результаты почти ежегодно проводимых учетных ихтиопланктонных и икорных съемок. В то же время запасы трески и камбал были относительно стабильными. Однако численность трески, размножающейся в районе юго-западного Сахалина, как показали данные учетной траловой съемки, в 2000 г. также заметно сократилась (более чем на 25 %).
У восточного Сахалина (рис. 11) запасы некогда многочисленных минтая и дальневосточной многопозвонковой песчанки из прол. Лаперу-за (Великанов, 1999; Зверькова, 1999) в 90-е гг. также характеризовались тенденцией значительного снижения, что в свете всего вышеска-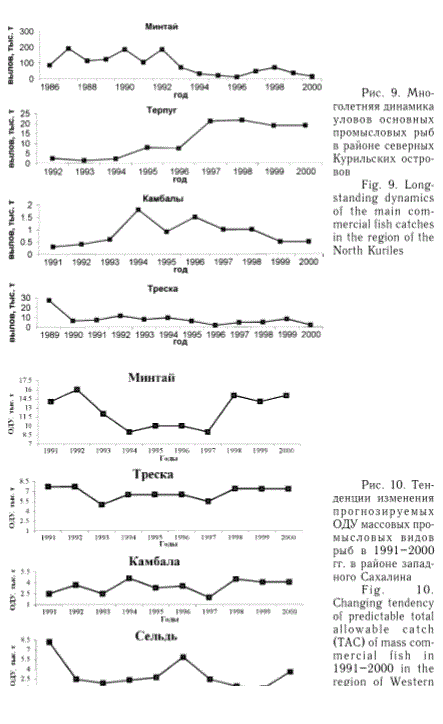
Рис. 9. Многолетняя динамика уловов основных промысловых рыб в районе северных Курильских островов
Fig. 9. Longstanding dynamics of the main commercial fish catches in the region of the North Kuriles
Рис. 10. Тенденции изменения прогнозируемых ОДУ массовых промысловых видов рыб в 1991-2000 гг. в районе западного Сахалина
Fig. 10. Changing tendency of predictable total allowable catch (TAC) of mass commercial fish in 1991-2000 in the region of Western Sakhalin
занного обусловлено ухудшением естественных условий воспроизводства этих видов. В это же время запасы камбал данного района существенно возросли. С одной стороны, это было связано с восстановлением популяции желтоперой камбалы в условиях запрета промысла и благоприятных фоновых условий ее воспроизводства, с другой, в меньшей мере, - с включением в ОДУ новых разведанных запасов камбал с дополнительным объемом изъятия более 1 тыс. т (звездчатая камбала северо-востока Сахалина, камбалы белобрюхая и Шренка зал. Анива).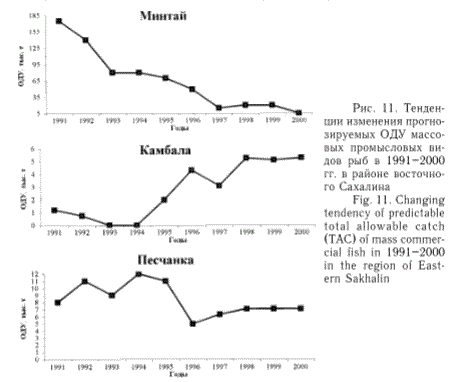
Рис. 11. Тенденции изменения прогнозируемых ОДУ массовых промысловых видов рыб в 1991-2000 гг. в районе восточного Сахалина
Fig. 11. Changing tendency of predictable total allowable catch (TAC) of mass commercial fish in 1991-2000 in the region of Eastern Sakhalin
У южных Курильских островов (рис. 12) в состоянии запасов минтая и трески в 90-е гг. отмечена тенденция снижения, запасы сайры и некоторых других рыб были относительно стабильны, а в запасах южного одноперого терпуга, анчоуса и ряда других во вторую половину или в конце 90-х гг. прослеживалась тенденция роста. Рекомендованные величины ОДУ по анчоусу, южному одноперому терпугу и некоторым другим видам рыб в указанный период лет были основаны на экспертных оценках их запасов. Однако, на мой взгляд, имеются достаточные основания считать, что тенденции изменения допустимых уловов анчоуса и южного терпуга, представленные на рис. 12, в общем соответствуют вектору реальной динамики запасов этих видов. В частности, об этом свидетельствует значительное увеличение численности японского анчоуса в летне-осенний период в южной части Охотского моря, наблюдавшееся во второй половине 90-х гг. при выполнении ежегодных эпипела-гических съемок, осуществляемых ТИНРО-центром (Шунтов и др., 1998а, б). Частично это подтверждается также обнаружением скоплений анчоуса в охотоморских водах Сахалина во второй половине 90-х гг. после 35-летнего перерыва, выявленных наблюдениями, проведенными СахНИ-
РО (Великанов, в печати). С начала 90-х гг. промысел анчоуса стал развиваться на охотоморском побережье Хоккайдо (Fishes and Marine inver-tebrates of Hokkaido ..., 1991). В 90-е гг. в южной части Охотского моря наблюдался также значительный рост численности южного одно-перого терпуга, биомасса молоди (сеголеток) которого в летний период, по данным В.П.Шунтова с соавторами (1998а, б), в отдельные годы достигала уровня 300 тыс. т. Аналогичного мнения о динамике запасов этого вида рыб в прибрежных япономорских и охотоморских водах о.
Хоккайдо в 90-е гг. Минтай придерживаются и
японские ихтиологи (Мицуширо Сано, устное сообщение). Еще более красноречиво об этом свидетельствует статистика японского вылова южного одноперо-го терпуга в северной части Хоккайдо. Так, с 1992 по 1998 г. уловы этой рыбы возросли втрое, достигнув в 1998 г. рекордной величины - более 170 тыс. т.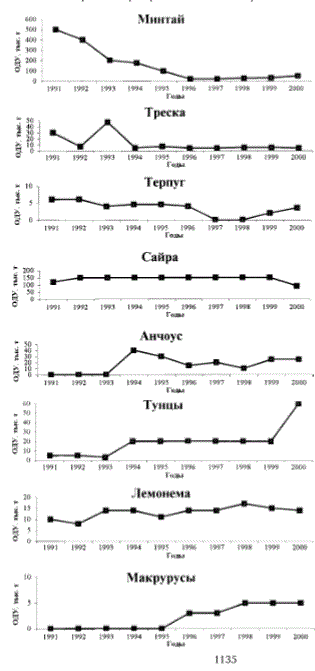
Рис. 12. Тенденции изменения прогнозируемых ОДУ массовых промысловых видов рыб в 1991-2000 гг. в районе южных Курильских островов
Fig. 12. Changing tendency of predictable total allowable catch (TAC) of mass commercial fish in 1991-2000 in the South Kuriles region
Что касается тенденций ОДУ по другим видам рыб (макрурусам, лемонеме, тунцам), то в условиях дефицита информации, возможно, они не отражают истиной динамики их запасов, но во всяком случае указывают на относительно удовлетворительное состояние этих сырьевых ресурсов. Однако для обоснованных выводов по возможностям современного промыслового освоения ресурсов этих рыб в водах южных Курильских островов необходима не только ретроспективная информация, но и перспективные специализированные поисковые исследования.
У северных Курильских островов (рис. 13) запасы минтая и трески в 90-е гг. характеризовались нестабильностью со снижением численности к концу десятилетия. Вместе с тем в многолетней динамике запасов северного одноперого терпуга и камбал наблюдалась тенденция возрастания численности.
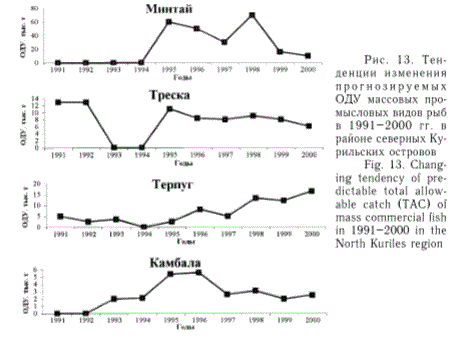
Рис. 13. Тенденции изменения прогнозируемых ОДУ массовых промысловых видов рыб в 1991-2000 гг. в районе северных Курильских островов
Fig. 13. Changing tendency of predictable total allowable catch (TAC) of mass commercial fish in 1991-2000 in the North Kuriles region
Таким образом, можно подчеркнуть, что в морских водах Сахалина и Курильских островов в настоящее время запасы минтая, трески, сельди, мойвы, песчанки характеризуются низким уровнем биомассы, а скопления сардины, скумбрии вообще отсутствуют. В противоположность, ресурсы таких рыб, как сайра, терпуги, анчоус, желтоперая камбала и некоторых других находятся в хорошем состоянии и могут обеспечить довольно высокий годовой улов. Конкретные цифры допустимых уловов наиболее значимых промысловых видов рыб на 2001 и 2002 гг. приведены в табл. 2. В целом современное состояние запасов морских промысловых рыб Сахалино-Курильского региона, включая пелагические, донные и глубоководные виды, позволяет изымать в 2001-2002 гг. только в режиме допустимого улова 480-540 тыс. т. Хотя приведенные суммарные величины прогнозируемого общего улова достаточно внушительны, тем не менее они подтверждают, что в начале нового столетия рыбопродуктивность морских акваторий рассматриваемого региона находится на относительно низком уровне. К примеру, еще в 80-е гг. предыдущего
века годовой вылов сардины иваси только в Охотском море был более 500 тыс. т (Zhigalin, Belayev, 1999), а в начале 30-х гг. того же столетия годовой улов сахалино-хоккайдской сельди, как известно, составлял около 1 млн т. Вместе с тем столь высокие уровни численности отдельных популяций рыб, обеспечившие такие годовые уловы, следует рассматривать скорее как исключение, чем норму. Во-первых, значительный по масштабам рост численности отмечен у очень ограниченного числа видов рыб, обладающих феноменально высоким экологическим потенциалом. В частности, как указывает В.П.Шунтов с соавторами (1997), полных экологических аналогов минтая и сардины иваси в дальневосточных морях нет. Во-вторых, масштабное увеличение численности вида (популяции) происходит в пределах одной природной циклики (клима-тоокеанологической эпохи) с продолжительностью 40-60 лет (Бышев и др., 1997; Шунтов и др., 1997). В связи с этим значительное увеличение рыбопродуктивности морских акваторий на определенном временном отрезке, например в одном столетии, будет происходить достаточно редко, не более 1-2 раз в 100 лет.
Таблица 2
Прогноз ОДУ основных промысловых видов рыб сахалино-курильского района в 2001-2002 гг., тыс. т
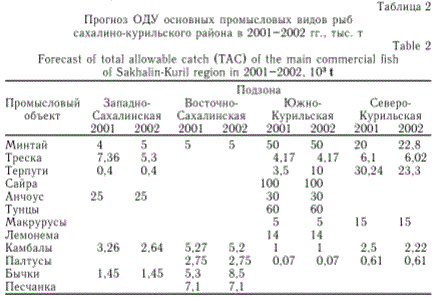
Учитывая вышесказанное, следует признать, что ожидать какого-либо существенного увеличения рыбных ресурсов в сахалино-курильском регионе (за счет роста численности популяций) в ближайшее время нет оснований. Правда, здесь необходимо сделать одну оговорку. В настоящее время рекомендованные величины ОДУ по южному одноперому терпугу, японскому анчоусу и некоторым другим видам рыб в сахали-но-курильском районе являются экспертными и весьма ориентировочными. Поэтому не исключено, что в случае более обоснованных уточнений их запасов общая величина ресурсов морских рыб на рассматриваемых акваториях в ближайшие несколько лет может возрасти. Вместе с тем следует иметь в виду, что в последнее десятилетие многие морские виды рыб рассматриваемого района очень слабо осваивались промыслом. В связи с этим все выявленные сырьевые ресурсы рыбных популяций представляют собой немалую перспективу для увеличения уловов в морских водах Сахалина и Курильских островов.
А.Я.Великанов (СахНИРО, г. Южно-Сахалинск)
Смотрите также






