Xимическая основа биопродуктивности Охотского моря
С точки зрения рыбохозяйственной деятельности человека под биопродуктивностью моря подразумевается прежде HCET) продуктивность промысловых o TexTOR B первую очередь ры бопродуктивность. На первый взгляд, между понятиями "химическая основа" и "рыбопродуктинность" никакой связи не существует. Дейстнительно, химические вещества в подавляющем большинстве случаев (за исключением острого дефицита кислорода и сероводородного заражения, загрязнения вредными веществами антропогенного происхождения) не оказывают непосредственного воздействия на рыбу. Однако, будучи частью морской экосистемы, рыба неизбежно становится зависимой от целого ряда веществ, которые используются на первом трофическом уровне для синтеза органики, жизненно важной для существования всех последующих трофических звеньев. Эта связь осуществляется через фитопланктон, который, синтезируя органику, извлекает из воды определенные минеральные вещества.
Большинство из них находится в морской воде в количествах, превышающих потребность в них фитопланктона. Содержание же в воде группы биогенных веществ зачастую бывает значительно ниже потребности в них фитопланктона, что лимитирует его развитие. Это минеральные соединения азота, фосфора, кремния, играющие важнейшую роль в метаболизме фитопланктона. В конечном итоге запас биогенных элементов в эвфотическом слое при прочих благоприятных условиях определяет величину продукции фитопланктона (первичную продукцию - ПП), на долю которой приходится более 90% синтезированного в море органического вещества. Именно фитопланктону принадлежит решающая роль в формировании кормовой базы различных гидробионтов, в том числе промысловых рыб.
Существование положительной связи между "химической основой" и "рыбопродуктивностью", таким образом, очевидно. Однако следует иметь в виду, что существует большое числотрофических уровней, по которым происходит передача энергии от фитопланктона к рыбе, которая находится почти на "вершине пищевой пирамиды моря". Кроме того, формирование рыбопродуктивности не ограничивается только трофическим фактором. Поэтому в данном случае речь не идет о каком-либо количественном выражении упомянутой зависимости, и задача ограничивается выявлением районов, потенциально наиболее благоприятных для создания хорошей кормовой базы для рыбы и других промысловых объектов, иными словами, районов, где наиболее высока обеспеченность фитопланктона биогенными злементами.
Материал, методнки
В настоящей работе использованы материалы экспедиционных наблюдений в Охотском море на РТМС "Млечный Путь" (март-апрель 1990 г.), НИС "Профессор Солдатов" (ноябрь-декабрь 1990 г.), НИС "Академик Александр Несмеянов" (июнь 1992 г. и июль 1993 г.). НИС "Академик М. Лаврентьев" (июль - август 1994 г.). Исследованиями охвачена большая часть моря, включающая глубоководную область, районы Курильских островов, Камчатского и Сахалинского шель
фов (рис.1). К сожалению, вне сферы внимания оказались такие высокопродуктивные районы, как заливы Шелихова и Терпения, а также северо- и западноохотоморский шельфы.
Использованы данные по содержанию в воде биогенных элементов - аммонийного, нитритного и нитратного азота, фосфора, кремния. Определение их концентраций (кроме аммонийного азота) проводилось на скоростном проточном автоанализаторе "Алпкем". В основе метода определения фосфора лежит реакция ортофосфатов с шестивалентным молибденом и трехвалентной сурьмой в кислой среде с образованием фосфосурьмяномолибденового комплекса, который затем восстанавливается аскорбиновой кислотой; оптическая плотность измеряется при длине волны 885 нм. Для определения нитритного азота используется его реакция с сульфаниламидом и N-1-нафтилэтилендиамином с образованием окрашенных диазосоединений, которые колориметрируются при длине волны 540 нм. При определении нитратного азота его предварительно восстанавливают до нитритного с помощью омедненного кадмия. Кремний определяется в кислой среде с молибдатом аммония и двухвалентным LTLLLLLLL L LLLLLL GLLL LLLLLLLLLLLLLL LLLLLS Для L. HIIREHIIII мешающего влияния ортофосфатов применяется винная кислота. Оптическая плотность измеряется при длине волны 820 нм. Определение аммонийного азота проводилось по методу Седжи-Солорзано с гипохлоритом и фенолом, оптическая плотность измерялась на спектрофотометре при длине волны 630 нм. Все перечисленные методики изложены в "Справочнике гидрохимика" (1991).
Обсужденне результатов
Рис.1 Схема расположения станций в Охотском море I - РТМС "Млечный Путь" (март - апрель 1990 г};
2- НИС "Профессор Солдатов" (ноябрь - декабрь 1990 Обеспеченность фитопланктона биогенными г.): 3 - НИС "Академик Александр Несмеянов" (июнь элементами оценивается по величине продукции, (uးဂိ'ိုမ်ိဳးနီ *ÇÑ ໃນ которую он может создать при использовании "iFዘ" " a . Лаврентье всего запаса биогенных элементов в эвфотичес
KOM OWL OER.
Основу этого запаса составляет их количество, которое формируется к началу вегетационного периода в результате осенне-зимнего конвективного перемешивания (рис.2, I). Кроме того, пополнение эвфотического слоя биогенными элементами из разных источников происходит и во время вегетационного периода. Наиболее важны те из них, которые ведут к повышению первоначального (зимнего) запаса биогенных элементов в эвфотическом слое и, как следствие, кувеличению биомассы фитопланктона, что, в свою очередь, способно вызвать возрастание продукции последующих звеньев пищевой цепи,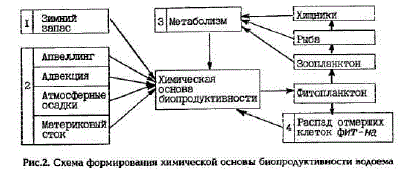
Рис.2. Схема формирования химической основы биопродуктивности водоема
Поступление биогенных элементов в эвфотический слой, осуществляемое в процессе подъема и горизонтальной адвекции вод, с материковым стоком и атмосферными осадками (рис. 2, 2) однозначно увеличивает их первоначальный запас.
В процессе метаболизма морских гидробионтов (рис.2, 3) видимого увеличения количества биогенных элементов не происходит. Однако их возврат в круговорот с продуктами метаболизма уже после эффективного использования фитопланктона морскими гидробионтами создает пред
посылки для дополнительного синтеза фитопланктоном органического вещества и, как следствие, для роста продукции на более высоких трофических уровнях. Кроме того, экскреция биогенных злементов может осуществляться мигрирующими гидробионтами, что сродни поступлению и минерализации аллохтонной органики. Таким образом, можно считать, что в процессе метаболизма запас биогенных элементов в эвфотическом слое фактически увеличивается.
Регенерация биогенных элементов при бактериальном распаде отмерших клеток фитопланктона, не востребованных фитофагами (рис. 2, 4), приводит лишь к восстановлению прежнего запаса питательных солей (и то не в полной мере - на 80-90%). Следовательно, при оценке химической основы биопродуктивности водоема только эту часть биогенных элементов следует исключить, так как синтезированная за их счет ПП не может внести никакого вклада в продукцию высших трофических звеньев. Хотя не следует приуменьшать значение регенерации, способствующей возврату биогенных элементов в воду и обеспечивающей, таким образом, более полное использование их запаса в эвфотическом слое.
Доля этой части продукции зависит от степени сбалансированности в отношениях фито- и зоопланктона. Она наибольшая, если максимум развития зоопланктона заметно отстает от максимума фитопланктона; в этом случае значительная часть клеток водорослей остается не использованной фитофагами и отмирает. Сближение во времени максимумов развития фито- и зоопланктона приводит к наиболее полному использованию продукции водорослей. В северной части Тикого океана выедание фитопланктона фитофагами начинается с очень небольшой задержкой (Heinrich, 1962), система фитопланктон - зоопланктон хорошо сбалансирована и фитопланктон потребляется почти полностью. Следовательно, можно считать, что в Охотском море основой создания кормовой базы для гидробионтов высших трофических уровней является ПП, созданная при использовании практически всего доступного фитопланктону количества биогенных элеменTOE
Зимний запас биогенных элементов формируется в результате осенне-зимнего конвективного перемешивания (рис.3). Чем оно глубже, тем больше коцентрация биогенных элементов в слое конвективного перемешивания. В нашем случае для характеристики зимнего запаса биогенных злементов использована их концентрация в остаточном холодном зимнем слое. Величина этого запаса биогенных элементов в эвфотическом слоев большой степени обусловлена его толщиной, которая варьирует в Охотском море в больших пределах: от 2 - 10 м в прибрежной зоне и областях подъема вод до 50 - 70 м в глубоководной части и местах антициклонических круговоротов (см.рис. З„г).
K началу вегетации фитопланктона запас азота, фосфора и кремния составил 250 - 2000 Mir-aT/M?, 25 - 150 MIT-aT/M? KA 500 - 4000 Mir-aT/M* cooTB eTciTBeH Ho (pHc.3, al-B). Ha 6owLLIIeň "HacTM Mcследованной акватории, главным образом в глубоководной области, зимний запас биогенных элементов был в следущих пределах - 1000 - 1250 мг-атN/м2, 60 - 100 мг-ат P/м2 и 1000 - 1500 Mr-ar Si/Mo. CaMbe Malle Benque (250 - 750 MT-ar N/Mo; 25 - 75 Mr-aTP/Mo; 500 - 1000 MT-aT Si/Mo) наблюдались в мелководных прибрежных зонах Сахалина и Камчатки, где наряду с относительно пониженной концентрацией азота, фосфора, кремния и толщина эвфотического слоя минимальна. Величины того же порядка характеризуют воды непосредственно над банкой Кашеварова и прикурильские воды, что здесь связано преимущественно с незначительной мощностью эвфотического слоя,
MaKCMMa AbHEile 3aniacEl 6MoreHHEIx 3AeMeHTOB (6onee 1250 MT-aT N/Mo, 6owee 100 Mr-ar P/Movi óomee 2000 Mir-aT Si/M?), обусловленные их высокой концентрацией и большой толщиной эвфотического слоя, приурочены, как правило, кaнтициклоническим вихрям (в том числе в районе материкового склона Сахалина и Камчатки) и к перифериям циклонических вихрей (в том числе в районе банки Кашеварова).
Приведенные величины определяют обеспеченность фитопланктона питательными солями к началу его вегетации. Закон минимума Либиха гласит, что "рострастений зависит от того элемента, который присутствует в самой низкой концентрации". Иными словами, применительно к морскому фитопланктону можно сказать, что его продукция лимитируется тем элементом, который находится в воде в относительном дефиците. Необходимо уточнить, что речь может идти только о так называемой новой продукции (причем минимальной ее величине), то есть продукции, синтезируемой фитопланктоном при утилизации лишь тех биогенных элементов, которые поступили в эвфотический слой извне, в данном случае, в результате осенне-зимнего конвективного перемешивания.
Стехиометрическое соотношение С:Si:N:Р = 106:23:16:1 для природных популяций фитопланкToHa [Redfield et al., 1963] CBMAleTem ELCTByeT, virTo III Mr C = 55 Si Mr-aT = 80 N MT-aT = 1272 Pмг-ат, а отношения Si/P; Si/N и N/P, в которых кремний, азот и фосфор потребляются фитопланктоном, равны 23, 1,4 и 16 соответственно. Анализ отношений биогенных злементов в верхнем слое воды показал, что перед началом вегетации фитопланктона на большей части акватории в относительном избытке находится кремний (Si/P более 23; Si/N более 1,4); в зоне шельфа же и материкового склона Сахалина и Камчатки, а также на северо-востоке, у входа в залив Шелихова - фосфор (Si/P менее 23; N/P менее 16)(рис.4). Азот же повсеместно был в относительном дефиците. Следовательно, на всей акватории Охотского моря азот будет использован фитопланктоном в первую очередь, и минимальная величина новой продукции будет эквивалентна зимнему запасу азота. Она составляет 10 - 150 г С/(м2 - год) (рис.5.a), и закономерности ее прост
ранственного распределения находятся в соответствии с распределением на исследованной Kватории величин зимнего запаса азота.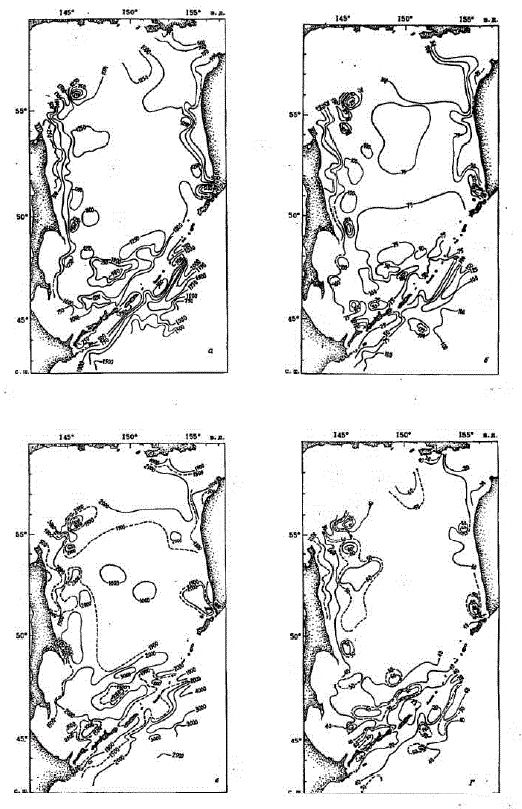
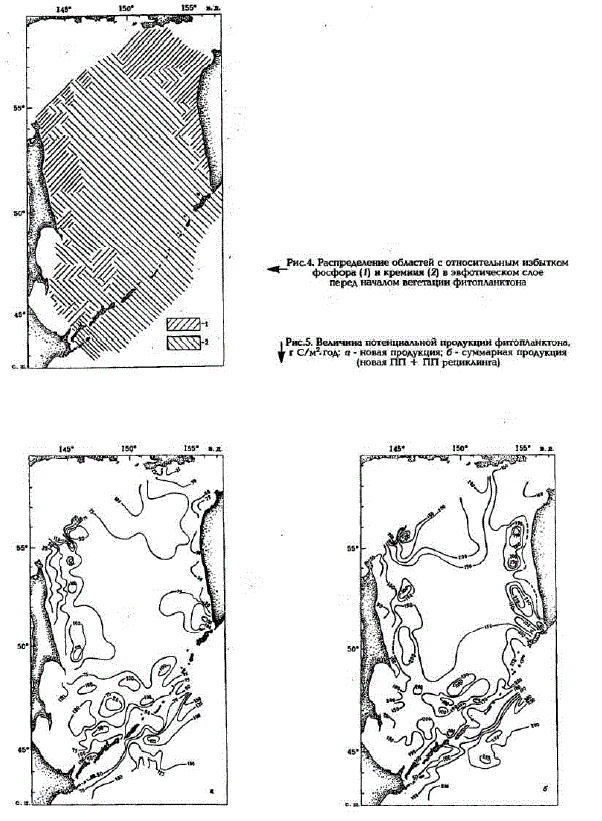
После исчерпания зимнего запаса азота повсюду, кроме областей апвеллинга, экосистема переходит на существование за счет рециклинга. Логично было бы предположить, что величина потенциальной суммарной продукции (новая ПП + ППрециклинга) должна быть эквивалентна количеству того элемента, который перед началом вегетации находился в относительном избытке. В действительности, независимо отсуществующей ситуации, она всегда обуславливается запасом кремния как наиболее консервативного из всех биогенных элементов. В то время как запасы азота и фосфора в течение вегетационного периода могут неоднократно оборачиваться в процессе рециклинга, значительная часть кремния из-за крайне медленного растворения скелетов и створок диатомей при высокой скорости их оседания сравнительно быстро выпадает из круговорота в верхнем слое воды. Кроме того, процесс растворения кремневых скелетов задерживается из-за наличия органических покровов и начинается лишь после того, как они разрушатся. Это приводит в конечном итоге к истощению эвфотического слоя кремнием и ограничивает дальнейшее развитие фитопланктона. Так, Уолш (Walsh, 1975), например, считает, что роль регенерации кремния в эвфотической зоне ничтожна. Однако А.Н.Богоявленский (1966), оценив величину потока взвешенной кремнекислоты из зоны фотосинтеза в глубинные воды, допустил, что 25 - 50% потребленного фитопланктоном кремния может все же возвратиться в воду в пределах эвфотического слоя. Причем можно предположить, что скорее всего это происходит в результате жизнедеятельности фитофагов, с продуктами метаболизма которых в воду поступают скелеты и панцири, уже лишенные органической оболочки, препятствующей их растворению.
При условии, что в течение вегетационного периода 50% зимнего запаса кремния вновь включается в оборот, суммарная потенциальная продукция составит 50 - 350 г С/(м2 - год) (рис.5.б). На значительной части исследованной акватории моря это максимально возможная величина продукции, которую может синтезировать фитопланктон на основе доступного ему запаса биогенных элементов. Исключение составляют области, на которые распространяется влияние прибрежного апвеллинга либо интенсивных циклонических вихрей, где происходит подъем глубинных вод обогащенных биогенными элементами, а также области, подверженные влиянию горизонтальной адвекции богатых биогенными элементами вод. В этих районах суммарная ПП значительно возрастает, причем за счет увеличения в ней доли новой продукции. Так, в районе банки Кашеварова, над которой в течение всего года существует интенсивный циклонический вихрь (Шунтов, 1985), при скорости подъема воды 101 см/с (Карпушин и др., 1996) и диаметре вихря около 80 км за сутки к поверхности поднимается около 430 км* воды, содержащей 22 мг-ат/м* азота, 2 мг-ат/м* фосфора и 40 мг-ат/м* кремния; эти богатые питательными солями воды распространяются к периферии вихря, создавая здесь резко выраженную градиентную зону. В целом такое поступление биогенных элементов обеспечивало бы за вегетационный период синтез около 1,8 - 108 т C, если бы все биогены утилизировались фитопланктоном.
Подобное обогащение эвфотического слоя питательными солями, хотя и в значительно меньшей степени, проявляется в областях материкового склона s" s". Сахалина и Камчатки, подверженных влиянию периодически возникающих прибрежных апвеллингов. Оценка суточной продукции при утилизации фитопланктоном биогенных элементов, поступающих в эвфотический слой в зоне прибрежных апвеллингов (Грузевич и др., 1996), показала, что она составляет 0,5% - 0,9 гC/(м2 - сут). В целом же в зоне материкового склона фитопланктон за вегетационный период может дополнительно синтезировать около 0,02-10°т С.
По-видимому, следует принять во внимание и то, что обеспеченность фитопланктона биогенными элементами в районе Курильских островов значительно улучшается за счет адвекции богатых питательными солями тихоокеанских Вод, а также в результате интенсивного вертикального перемешивания их в проливах Курильской гряды.
В пересчете на всю исследованную площадь моря продукция фитопланктона, которую он может создать на основедоступного ему количества биогенных элементов, составляет 4.0 - 10°т C в год, в ней на долю их зимнего запаса приходится 22 - 108 тС.
Следует признать, что итоговая величина продукции несколько занижена, так как в настоящих расчетах не вполной мере учтено пополнение эвфотического слоя биогенными элементами за счет подъема глубинных вод и совсем не поддается учету поступление их за счет горизонтальной адвекции. Крометого, не принятово внимание влияние материкового стока и атмосферных осадков.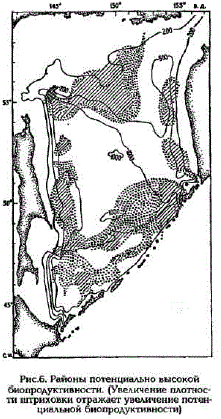
Подводя итог полученным результатам, можно констатировать, что благоприятные условия для формирования высокой биопродуктивности существуют на значительной части исследованной акватории (рис.6). На общем фоне выделяют- Рис.б. Районы потенциально высокой ся районы над материковым склоном Сахалина и Камчатки, биопродуктивности. (Увеличение плотности штриховки отражает увеличение потенгде в поле течений наличествуют сопряженные вихревые об- циальной биопродуктивности)
разования район, примыкающий к северно-восточной части Курильский гряды, куда через проливы Крузенштерна и Четвертый Курильский поступают богатые биогенными элементами тихоокеанские воды; районы на северной границе Курильской котловины и в центральной части ее, в области антициклонического круговорота.
Особого внимания заслуживает район банки Кашеварова, где количество биогенных элементов, поступающих в центре циклонического вихря в поверхностные воды, практически равноценно суммарному запасу их в эвфотическом слое на всей исследованной акватории моря.
Н.В.Аржанович, В.Л.Зубаревич
Смотрите также






