Первичная переработка в Охотском море
Охотское море является одним из главных рыбопромысловых регионов России с годовым выловом около 1,5 млн. т. (Шунтов, 1984: Шунтов и Дулепова, 1991). Данные о высокой биомассезоопланктона (Кун, 1957; Лубны-Герцык, 1959; Маркина и Чернявский, 1984), фитопланктона (Смирнова, 1959) и зообентоса (Савилов, 1961, Маркина и Чернявский, 1981) подтверждают представление о высокой биологической продуктивности этого субарктического бассейна и, соответственно, о высокой первичной продукции в его пелагиали. Последний вывод, хотя и не был обоснован прямыми определениями активности фотосинтеза, нашел отражение в картах распределения первичной продукции в северной части Тихого океана (Кобленц-Мишке и др., 1970, Sanger, 1972). Путем экстраполяции результатов ее измерения в сходном субарктическом бассейне Берингова моря вероятный уровень первичной продукции в Охотском море оценивали в 0,3-0,6 гC/(м* - сут). Единичные прямые определения ее кислородным методом склянок в водах мелководных заливов Анива и Терпения дали значения в пределах 0,1-0,5 мг C./(л" сут) или 0,03 - 0,2 гC/м? (Сафонова, 1990). Косвенные расчеты первичной дроукции по убыли концентрации фосфатов в толще водыдали величины продукции 0,2-0,3 г С/м" в летний период (Сметанин, 1956). Вполне очевидно, что для получения достаточно надежных оценок этого важного показателя биопродуктивности Охотского моря необходимы были прямые определения с помощью современной методики. Такие определения были выполнены нами в июле-августе 1992 г. с помощью радиоуглеродного метода в рейсе НИС "Академик Александр Несмеянов",
Материал и методнка
Измерения первичной продукции были выполнены на 40 станциях основных разрезов через
центральную часть моря и на станциях трех шельфовых полигонов: Амурского - в Сахалинском заливе, Пильтунского и Луньского - у Северо-Восточного побережья Сахалина (рис.1).
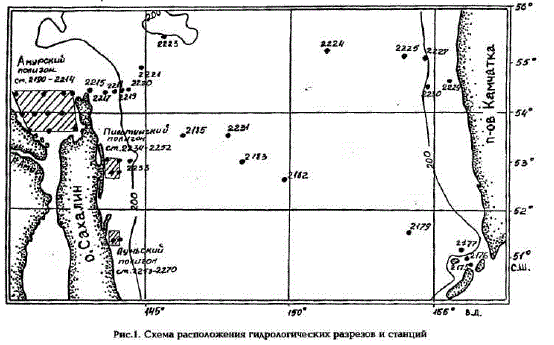
Рис.1. Схема расположения гидрологических разрезов и станций
Всего было выполнено около 500 индивидуальных определений интенсивности фотосинтеза. Пробы для анализа отбирали из пластмассовых батометров зонда "Нил Браун". Горизонты для отбора проб в толще воды выбирали с учетом результатов предварительного зондирования распределения температуры, солености и флуоресценции хлорофилла. Первичную продукцию измеряли с помощью радиоуглеродного метода в модификации автора (Сорокин, 1960, 1987). Для опре: делений пользовались склянками объемом 300 мл. Абсолютные величины фотосинтеза (Рs, гC/м* за сутки) измеряли в пробах воды из верхнего слоя в двойном повторе с одним темновым контролем при экспозиции склянок на палубе в условиях естественного освещения и допустимых колебаний температуры. Специальными опытами было показано, что колебания температуры в таких пределах мало влияют на результаты измерений. Продолжительность экспозиции склянок в аквариуме составляла 2 - 3 ч, если пробы были отобраны между 8.00 и 14.00 часами, и 4 - 5 ч, если они отбирались позже или раньше. Пробы, отобранные вечером после 18.00 часов или ночью, выдерживали до утра в палубном аквариуме в двухлитровых склянках. В 7 часов утра их разливали в склянки для экспозиции. Для вычисления суточных величин первичной продукции (P) по результатам измерений ее в кратковременных опытах (P) использовали коэффициенты К которые соответствовали P/P определенным для различных периодов светового дня. Величины коэффициентов K для разных отрезков светового дня были получены путем измерения суточного хода фотосинтеза (рис.2). Их значения приведены в табл.1. Величины интегральной продукции под 1 м2 вычисляли, пользуясь измеренными на станциях величинами P и кривыми OITH CHIBrthющими изменение скорости фотосинтеза в толще воды с глубиной. Коэффициенты К вычисляли перемножением световых коэффициентов К. и коэффициентов. К характеризующих относительное распределение активного фотосинтеза в толще воды (Сорокйн, 1960). Примеры соответствующих кривых K и К приводятся на рис.3.
R Таблшца І. Коэффициенты (KI) для расчета 500 первичной продукции заполный световой день
с использованием ее часовых величин, 4DO измеренных кратковременных OTTO
на разных отрезках светлого периода суток
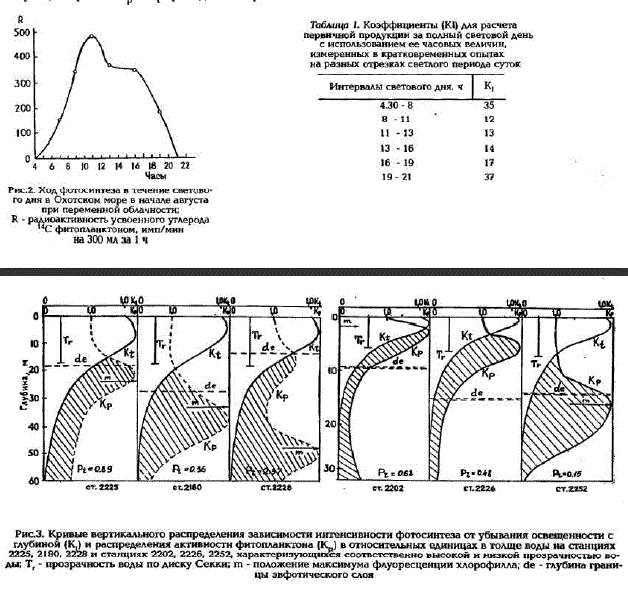
Рис. 3. Кривые вертикального распределения зависимости интенсивности фотосинтеза от убывания освещенности с глубиной (К) и распределения активностн фитопланктона (К) в относительных единицах в толще воды на станциях 2225, 21 80, ה, 8ללל станциях 2202, 2226, 2252, характеризующихся соответственно высокой и низкой прозрачностью воды; T, - прозрачностъ воды по диску Секки; пn - положение максимума флуоресценции хлорофилла; de - глубина границы эвфотического слоя
Результаты
Результаты измерений первичной продукции на станциях основных разрезов приводятся в табл.2. Они показывают, что в открытых Päಂನ್ಡು моря на большей части станций в поверхностном слое воды она превышала 60 мг С/(м* - сут) и была более 0,4 0,5 г С/(м2 - сут) в толще воды. Пределы ее колебаний на станциях составляли от 2 до 267 мг C/м3 в поверхностном слое и от 0,03 до 2.73 г С/(м* - сут) в толще воды. Максимальные величины первичной продукции были отмечены на шельфе в районе материкового склона о. Парамушир и п-ова Камчатка на ст. 2175-76, 2228
Таблица 2. Первичная продукция за сутки в поверхностном слое воды (Ps, мг Clм") нее интегральные величины в толще воды (Pt.т См") на станциях основиых разрезов в открытых районах Охотского моря и на шельфовых полнгоиах у о.Сахалнн
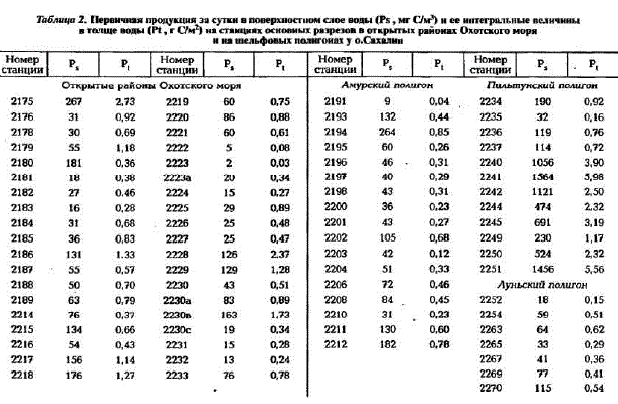
Самые низкие величины первичной продукции неожиданно были обнаружены над банкой Кашеварована ст. 2222-23 (0,03 - 0,08 гC/м"). Столь низкая первичная продукция на этих станциях при высокой концентрации биогенов была, по-видимому, следствием слабой вертикальной устойчивости вод, обусловленной воздействием на течения подводной воз
BEIHHOT
Несмотря на то что наши измерения первичной продукции приходились на время сезонного минимума фитопланктона, характерного для летнего периода в субарктических бассейнах (Гейнрих, 1961), средняя ее величина на станциях основных разрезов оказалась весьма высокой - около 60 мг C/м3 и около 0,80 гC/(м2 - сут) (см.табл.2). Этот феномен объясняют в значительной мере особенности вертикального распределения гидролого-гидрохимических характеристики активного фитопланктона.
В рассматриваемый период верхняя граница термоклина (пикноклина) находилась в пределах освещенного слоя на глубинах 15 - 25 м, при глубине нижней границы последнего 30-60 м. С положением термоклина совпадает и градиентный слой солености. Поэтому оседающее сообщество диатомейскапливалось и поддерживалось у верхней границы термоклина (пикноклина) в пределах освещенной зоны, куда осуществлялся приток биогенов из градиентной зоны (рис. 4, а, б).
На ст. 2228-30 упобережья Камчатки высокая первичная продукция имела место в условиях, когда основная часть весеннего сообществадиатомей уже осела к нижней границе освещенного слоя (см. рис.4,в, ст. 2229), а у вторичного термоклина, возникшего вверхнем перемешанном слое на глубине 12 - 15 м, уже сформировалось новое летнее сообщество диатомей с доминированием видовChaltoceros и Rhyzosolenia. Таже картина опускания активного фитопланктона книжней границе освещенного слоя прослеживается в районе материкового склона в юго-восточной части моря (см. рис.4, г. ст. 2179).
Наст. 2175 на шельфе о. Парамушир (Курильские о-ва) сообщество фитопланктона находилось в переходной фазе от весны к лету. Граница пикноклина оставалась здесь еще на глубине 1015 м, а фитопланктон был сконцентрирован в слое 5 - 10 м с максимумом на глубине 10 м при глубине залегания границы освещенного слоя 30 м. Та же ситуация была отмечена на ст. 22172220 на шельфе и над склоном у Северного Сахалина (см. рис.4,9,e).
Результаты измерений первичной продукции на трех шельфовых полигонах у о. Сахалин приведены в табл.2.
На станциях Амурского полигона в Сахалинском заливе, над глубинами от 15 до 60 м, высокая активность фотосинтеза в поверхностном слое воды - от 60 до 260 мг С/(м* - сут) была зарегистрирована в полосе распространения распресненных поверхностных вод из эстуария Амура. Наличие резкого пикноклина на небольшой глубине поддерживает здесь сообщество фитопланктона в пределах неглубокой эвфотической зоны (12 - 20 м) вследствие большой мутности и малой прозрачности воды (см. рис.4,ж, ст. 2202). Интегральные величины первичной продукции в этой полосе были умеренными, составляя 0,50- 0,86 г. С/(м2- сут). На других станциях полигона продукция фитопланктона была ниже: 0,2-0,4 г С/м*.
На Пильтунском полигоне, над глубинами от 16 до 260 м, даже в начале августа, когда проводились наши наблюдения, сообщество фитопланктона все еще пребывало в весенней стадии cезонной сукцессии при температуре воды у поверхности 4,5 - 6,0°С и глубине верхней границы пикноклина 8 - 15 м (см. рис.4,ж, ст. 2240, 4а, ст. 2241; 4з, ст. 2251). В прибрежной же и центральной частях полигона сохранялось пятно типичного массового цветения диатомовых водорослей с биомассой фитопланктона до 15 г/м3. Первичная продукция на станциях в этом пятне была очень высокой, близкой к ее верхнему пределу для природных сообществ фитопланктона открытого моря: до 156 мг С/(м* - сут) в поверхностном слое и до 6 гC/м2 - в толще воды (см.табл.2). У восточного края полигона, вне пятна цветения, основная масса водорослей уже осела к нижней границе эвфотической зоны и их продукция снизилась (см. рис.4,з, ст. 2242).
В пятне остаточного весеннего цветения диатомей основная масса фитопланктона была представлена талассиозирой Th. пordenskioldiі, считающейся холодолюбивым видом диатомей [Смирнова, 1959: Семина, 1974). После прогрева воды и перехода к летней фазе сукцессии основная масса этих диатомей оседает к нижней границе зоны фотосинтеза и затем вытесняется представителями родов Chaetoceros, Nitzschia, Ditilшп, Rhizosolenia. На станциях у восточной окраины Пильтунского полигона Talassiosira nordenskioldiї оставалась доминирующим видом и при температуре 8 - 9"C. В связи с этим возникли вопросы о том, насколько данный вид диатомей является холодолюбивым и в какой степени его вытеснение другими видами водорослей в начале летнего прогрева действительно связано с этим его свойством. Мы попытались решить эти вопросы, измерив интенсивность фотосинтеза при разных температурах в пробах воды, взятых в пятне цветения, где Т. погdenskioldiі составляла более 90% всей биомассы. Результаты опыта, представленные в табл.3, показывают, что интенсивность фотосинтеза водорослей в пробе не только не тормозится при температуре выше 5°С, но растет до температуры 14"C, что заставляет усомнится в истинной психрофильности данного вида диатомей.
На Луньском полигоне, располагавшемся на 1" широты южнее, мы наблюдали раннелетнюю фазу сукцессии фитопланктона, когда основная масса водорослей уже осела к нижней границе освещенной зоны в результате прогрева и опускания пикноклина. Максимумы концентрации фитопланктона находились на глубинах 20 - 30 м, и основная его масса испытывала недостаток света (см. рис.4, и). Тем не менее первичная продукция на станциях полигона была все еще достаточно высокой - до 115 мг С/(м* - сут) у поверхности и до 0,62 гC/м2 в толще воды.
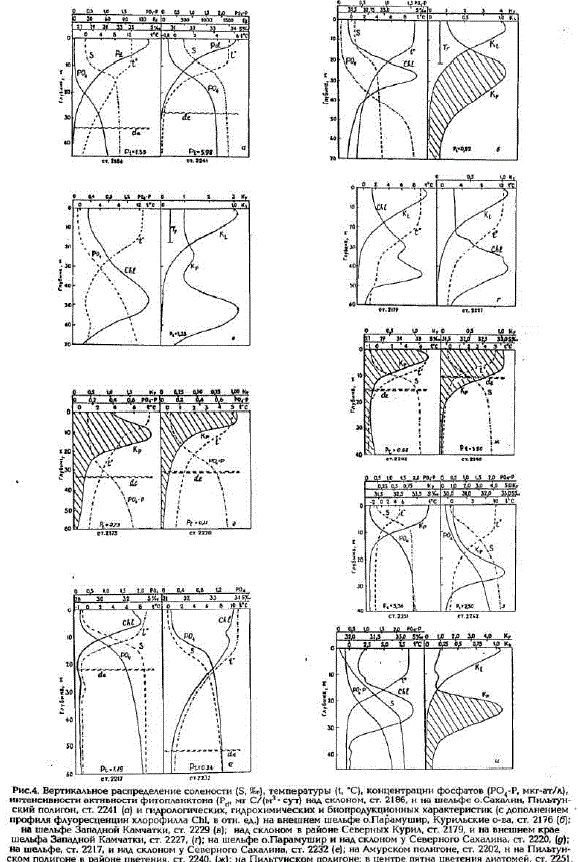
Рис.4. Вертикальное распределение солености (S. it), температуры (t, "С), концентрацни фосфатов (РС-Р, мкг-ат/л), интенсивности актнвности фитопланктона (P. мг С/(м* - сут) над склоном, ст. 21В6 и на шельфе о Сахалин, Пильтунский полигон, ст. 2241 (а) и гидрологических гидрохимических и биопродукционных характеристик (с дополнением профиля флуоресценцин хлорофилла Chl, в отн. ед.) на внешнем шельфе о.Парамушир, Курильские о-ва, ст. 2176 (б): нашельфе Западной Камчатки, ст. 2229 (в) над склоном в районе Северных Курил, ст. 2179, и на внешнем крае шельфа Западной Камчатки, ст. 2227, (г), на шельфе о.Парамушир и над склоном у Северного Сахалина, ст. 2220, (g); на шельфе, ст. 2217, и над склоном у Северного Сакалина, ст. 2232(e); на Амурском полигоне, ст. 2202. н на Пильтунском полигоне в районе цветения, ст. 2240, (ж) на Пильтунском полигоне: в центре пятна цветения диатомей, ст. 2251, и на его периферии, ст. 2242, (5): на Луньском полигоне, ст. 2263, (и). P - первичная продукция, г Č/M?: de . глубина эвфотической зоны
На многих станциях в центральной части моря сообщества диатомового фитопланктона B peзультате весеннего, летнего прогрева и опускания пикноклина уже осели к нижнеигранице освещенной зоны и образовали максимумы концентрации на глубинах 40-50 м (см. рис.3; 4,в,г). Эти максимумы прослеживались на профилях флуоресценции хлорофилла и на кривых распределения активного фитопланктона К. Освещенность на этих глубинах составляла всего 3 - 5% ФАР. Хорошее совпадение обоих MaKdßiMyMoB показывает, что фитопланктон в них находился в активном состоянии и интенсивно фотосинтезировал при достаточной освещенности. В связи с этим возникает вопрос о его адаптированности к слабому свету. В последнем случае следовало ожидать, что он может давать достаточно высокую продукцию, находясь в слое максимума у нижнеи границы освещенной зоны. Такая световая адаптация фитопланктона в слоях его глубинных максимумов отмечалась ранее в водах тропических и умеренных широт. Однако опыты с экспозицией склянок с пробами, взятыми из слоя такого максимума, не подтвердили этого предположения (табл.4). По-видимому, оседание продуцирующих сообществ диатомей в субарктических водах в начале лета происходит настолько быстро, что они не успевают адаптироваться к пониженной освещенности. Испытывая недостаток света, они постепенно отмирают и замещаются видами диатомей, обладающими большей способностью удерживаться в толще воды при малых градиентах плотности, или жгутиковыми формами водорослей.
Таблица З. Влияние температуры наннтенсивность Таблица 4. Влняине освещенности фотосинтеза в пробах воды, отобрапных в пятне на скорость фотосиптева в пробал, остаточного весеншето цветения таласснознры отобранных с поверхностин на слоя T. 
Обсуждение результатов нсследования
По показателям рыбопродуктивности, по обилию фитопланктона, зоопланктона и бентоса Охотское море давно уже относили к самым продуктивным морским бассейнам (Зенкевич, 1963; Маркина и Чернявский, 1984; Шунтов, 1985). В период весеннего цветения диатомей биомасса фитопланктона может достигать 5 - 15 мг C/м* даже в открытых его районах (Смирнова, 1959; Микаэлян и др., 1995).
Результаты наших определений показали, что первичная продукция в этом бассейне также достаточно высока даже в период летнего прогрева и сезонного минимума фитопланктона в июлеавгусте (см.табл.2). В табл.5 представлены ее величины, осредненные по основным районам
Таблица 5. Средние велпчнны первичной продукции, содержанне хлорофилланбногенов в различпых райопях Охотского моря"
Первичная продукция Биогены
Р и Р - первичная продукция за сутки: ChI - содержание хлорофилла в воде; P/B - удельная продукция фитопланктона за сутки A - ассимиляционное число хлорофилла, мг СУмг клорофилла "а" в час: N - число станций, для которых рассчитывались средние величины
пелагиали. Как видно из этой таблицы, первичная продукция оставалась сравнительно высокой даже в наименее продуктивных открытых районах, испытывающих сильное влияние обедненных планктоном и биогенами вод тихоокеанского происхождения (Чернявский, 1981). Средняя еевеличина в поверхностном слое составляла здесь 44 мг С/(м* - сут). В северо-западной части исследованной акватории, занятой преимущественно собственно охотоморскими водами, средняя продукция была выше 84 мг C/м". На шельфе о.Сахалин (вне пятен цветения диатомей) она составляла 90 - 91 мг С/м. (см. табл.5), но резко возрастала в зоне цветения на Пильтунском полигонедо 984 мг C/м3 при максимуме 1500 мг C/м", что соответствует ее уровню в районах интенсивного прибрежного апвеллинга тропической зоны океана. Интегральные величины первичной продукции 3-6 г С/(м*- сут), наблюдавшиеся в пятне цветения, могут служитъ мерой ее уровня в шериод массового весеннего развития диатомей, охватывающего всю акваторию моря после таяния льдов (Гейнрих, 1961; Смирнова, 1959). Разрушение и исчезновение ледяного покрова происходит неравномерно, и пятна цветения можно встретить (см.табл.2) даже в начале августа у Восточного побережья Сахалина. В наблюдавшемся нами пятне остаточного весеннего цветения преобладала Т. погdenskioldii, формировавшая до 90% суммарной биомассы. В прилегавших к пятну со стороны склона более теплых водах масса водорослей остаточного весеннего цветения диатомей уже осела к границе термоклина или исчезла и была замещена в слое максимума фитопланктона другими диатомеями из родов Chaetoceros, Ditilшm, Rhyzosolenia. Cмену видов диатомей, формирующих весеннее цветение в субарктических морях, и в частности, исчезновение наиболее массового вида Т.погdenskioldiі, связывают предположительно с его психрофильностью [Смирнова, 1959). Однако эксперимент (см.табл.4) не подтвердил предполагаемую психрофильность этого вида. Интенсивность фотосинтеза в пробах воды из пятна цветения, в которых доминировала эта диатомея, не подавлялась даже при температуре 14"С. Одна из возможных причин вытеснения Т. пordensioldiі при прогреве воды может быть связана с тем, что она имеет массивные гладкие створки и быстрее оседает по сравнению с такими водорослями, как Chaetoceros, более приспособленными для парения в толще воды. Возможно, что основным условием для массового развития талассиозиры является резкий пикноклин в самом верхнем, хорошо освещенном слое воды на глубине 3 - 10 м. Именно такие условия существуют в субарктических морях после весеннего таяния льдов и начинающегося прогрева верхнего слоя, когда температура его достигает 3 - 5°С (см. рис.4,ж.з). В этом случае основная массадиатомей удерживается над пикноклином. Будучи сконцентрирована в тонком верхнем распресненном слое, эта масса диатомей задерживает свет, замедляя тем самым опускание термоклина в глубину по мере прогрева верхнего слоя. В конце концов под влиянием прогрева, при температуре В - 10"С у поверхности, граница термоклина начинает опускаться, и с HeM погружается в глубину масса талассиозиры, сначала формируя максимумы у нижней границы освещенной зоны (см. рис.4,з,и). Постепенно отмирая из-за недостатка света, популяция этого вида диатомей заменяется видами Chaetoceros, Rhyzosolenia, Ditilшлп, Cosсіпоdiscus и жгутиковыми формами.
Физиологическая активность сообществ фитопланктона была весьма высокой. Часовые ассимиляционные числа хлорофилла вверхнем слое воды Охотского моря были в среднем вдвое выше, чем в летний период вводах умеренных широт: 5-9 мг C/мг хлорофилла "а", против 3 - 5 мг C/мг хлорофилла в час соответственно (Кобленц-Мишке и Ведерников, 1977). Удельная продукция (P/B-коэффициенты) в верхнем слое воды Охотского моря была также достаточно высокой - выше единицы за сутки (см. табл. 5). Эти данные указывают на достаточно хорошую обеспеченность сообществ фитопланктона биогенами даже в верхнем перемешанном слоев период сезонного минимума его развития. Абсолютная концентрация фосфатов варьировала в этом слоев широких пределах - от 0,01 до 0,80 мкг-ат/л (см.табл.5, рис. 4, 5).
Ниже границы термоклина концентрация фосфатов быстро нарастала до 1,5-2 мкг-ат/л. Отсутствие корреляции между величинами первичной продукции и концентрацией фосфатов в верхнем слое воды (рис.5) указывает на высокую интенсивность потоков биогенов в пелагиали Охот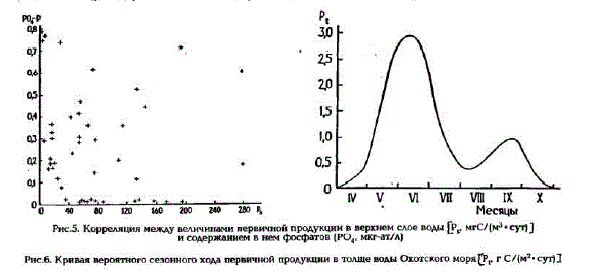
Рис.5. Корреляция между величинами первичной продукции в верхнем слое воды P MrC/Mo-cyr) и содержанием в нем фосфатов (PC, мкг-ат/л)
Рис.б. Кривая вероятного сезонного хода первичной продукции в толще воды Охотского моряUР. r C/(M? • cyrI
ского моря в летний период. Их поступление в верхний слой может осуществляться, в частности, за счет подъема вод в циклонических мезомасштабных вихрях, образующихся при взаимодействии разнонаправленных течений или вследствие воздействия на течения сложного рельефа дна (Чернявский и др., 1981). В результате такого взаимодействия течений с рельефом формируются несколько зон локальных апвеллингов у побережья Камчатки и Сахалина (Маркина и Чернявский, 1984). Другой источник постоянного пополнения биогенов - их регенерация из органических соединений. Расчетное время оборота минерального фосфора в верхнем перемешанном слое Охотского моря составляет 3-4 сут в открытом море и околосуток у Западного побережья о.Парамушир и п-ова Камчатка. На мелководных полигонах у о. Сахалин расчетное время оборота фосфора составило 4-6 сут (см.табл. 5). Содержание минерального азота (NO3 + NH-N) варьировало вверхнем слое в пределах 1 - 3 мкг-ат/л. Эти величины также слабо коррелируют с высокой в среднем первичной продукцией, что лишний раз свидетельствует о том, что не абсолютная концентрация биогенов, а скорее интенсивность их поступления в зону фотосинтеза определяет продуктивность и физиологическую активность сообществ фитопланктона встратифицированном морском бассейне (Sогоkin, 1985].
Отношение интегральной первичной продукции в толще воды (P) к таковой в поверхностном ее слое (Р) в прозрачных водах северо-восточных районов центральной части моря составляло в среднем около 15 (см.табл. 5). В более продуктивных и менее прозрачных водах северо-западной части исследованной акватории отношение P/P было около 9 и на прибрежных полигонах у Сахалина - 4-6. Эти соотношения могут быть использованы в ходе последующих измерений первичной продукции в Охотском море в летний период.
Учитывая данные, которые дают представление об уровне первичной продукции в зонах весеннего цветения диатомей и в районах моря, где уже наступила летняя фаза сезонной сукцессии фитопланктона (см.табл.2), а также принимая во внимание сезонный ход развития фитопланктона в субарктических бассейнах (Гейнрих, 1961), мы построили кривую вероятного сезонного хода первичной продукции в Охотском море (рис.6). Эту кривую мы использовали для расчетов годовой дРодукции фитопланктона. Согласно расчетам, она должна ориентировочно составлять 220 г. С/(м2- год) или 340-105 т углерода в расчете на всю акваторию моря. Эта величина вдвое ниже ранее сделанной на основе косвенных расчетов оценки (Маркина и Чернявский, 1984; Маркина, 1990].
Полученные нами расчетные величины годовой первичной продукции хорошо соотносятся с расчетной величиной годового рациона мирных гидробионтов (зоопланктона и зообентоса) в Охотском море, составляющего, согласно В.П.Шунтову и Н.И.Дулеповой (1991), 350 млн. тутлерода.
Ю.И.Сорокин (ИО РАН, Южное отделение)
Смотрите также






