Биохимический состав взвешеного и растворенного органического вещества в Охотском море
Изучение биохимического состава органического вещества (ОВ) позволяет, с одной стороны, судить о происхождении ОВ, а с другой стороны, проследить за его трансформацией в морских экосистемах. К настоящему времени выделены тысячи различных соединений, образующих морcKoe OB Lee, Wakeham, 1988; Lee, Henrichs, 1993), oAHako B. MopCKHx 6PoxMMMecKMx AccaeaoBaниях особое внимание уделяется таким главным биохимическим компонентам OB, как углеводы, белки, липиды и нуклеиновые кислоты (HK).
Основным продуктом первичного продуцирования являются углеводы, которые затем преобразуются как в результате клеточного метаболизма, так и метаболизма всей экосистемы водоема. Отмечена сильная корреляция между количеством взвешенных углеводов и биомассой фитошланктона [Marshall, Orг, 1962: Eberlein, Brockпmann, 1986; Liebezeit, 1987], а также между интенсивностью первичного продуцирования и количеством растворенных углеводов (Sellner, 1981). Таким образом, изучение пространственно-временной изменчивости количества углеводов в морских экосистемах дает представление об интенсивности первичного продуцирования и изменении запасов вещества и энергии данной экосистемы.
Концентрация белка, особенно во взвеси, дает представление о количестве гетеротрофных микроорганизмов, которые являются основными трансформаторами как автохтонного, так и аллохтонного ОВ (Cho, Azam, 1988; 1990). По колебанию его концентраций можно не только проследить за изменением биомассы микропланктона, но также оценить запасы органического азота в изучаемой экосистеме (Tanoue, 1995).
Кроме белка, в содержание взвешенного и растворенного органического азота большой вклад вносят НК. Они являются важными поставщиками не только органического азота, но и органического фосфора. В морской среде HK в основном продуцируют активно растущие микроорганизмы и усваивают гетеротрофные бактерии, гидролизующие их внутриклеточными и внеклеточными нуклеазами [Karl, Bailiff, 1989]. Оборот НК в экосистеме водоема происходит очень быстро (в течение 3 - 6 ч.), поэтому, несмотря на небольшое относительное содержание в ОВ, их вклад в круговоротазота и фосфора очень велик.
Липиды, благодаря своей большой химической устойчивости, могут служить трассерами поступления OB в данную акваторию и распространения в ней [Gerin, Goutк, 1994].
Следует отметить, что важным биохимическим показателем является не столько концентрация какого-либо индивидуального компонента, а и его относительное содержание в растворенном и взвешенном ОВ. Соотношения основных биохимических соединений РОВ и ВОВ позволяют судить о происхождении, путях трансформации и пищевой ценности ОВ для высших трофических уровней морской экосистемы.
К сожалению, в литературе нет данных о биохимическом составе РОВ и ВОВ в Охотском MOpe.
В течение трех лет (1992 - 1994 гг.), в летний период мы проводили исследование распределения ОВ и его основных биохимических компонентов в различных районах Охотского моря: в глубоководной его части, в области шельфа и материкового склона Камчатки и Восточного Сахалина Сахалинском заливе и в Прикурильском районе. В 1993 г. впервые были выполнены работы по определению биохимическиго состава ОВ в поверхностном микрослое (ПМС). В данной статье проведен анализ распределения растворенных и взвешенных углеводов, белков, липидов и НК.
Матерпалы и методы
Материалы были получены в 21-м и в 24-м рейсах НИС "Академик Александр Несмеянов" в августе 1992 г. и июле-августе 1993 г., а также в 21-м рейсе НИС "Академик Лаврентьев" в июле - августе 1994 г. (рис. 1).
Отбор проб воды для биохимического анализа проводили из пластиковых батометров зондирующего комплекса Нейл-Браун. Воду из батометров сливали в полиэтиленовые канистры через газ с ячеей 60 мкм для отделения зоопланктона и крупного фитопланктона. Отбор проб воды в ПMC описан в статье С.А.Соколовой с соавторами (1995). Взвесь собирали на стекловолокнистые фильтры CF/F (0.5-0.2 мкм), предварительно прокаленные при 480°С. Из собранной взвеси вместе с фильтром готовили гомогенаты вручном стеклянном гомогенизаторе. Затем в воде после фильтрации и в полученных гомогенатах определяли концентрации основных биохимических компонентов РОВ и ВОВ (Справочник., 1991).
Определение растворенного белка и полипептидов проводили после их сорбции из морской воды на стекловолокнистых фильтрах CF/F и после окраски Кумасси R-250. Максимум поглощения окрашенного комплекса - 590 нм.
Взвешенный белок определяли по методу Лоури.
Липиды из морской воды экстрагировали смесью хлороформа с этанолом и метанолом в соотношении 2:1:1 по объему. Взвешенные липиды экстрагировали этой же смесью. В экстрактахлипиды определяли по образованию окраски в результате сульфофосфованилиновой реакции.
Рис.1. Схема расположения биохимических станций
Максимум поглощения окрашенного комплекса - при длине волны 530 нм. Контролем при определении взвешенных липидов служил органический экстракт из суспензии бланкового фильтра CF/F; контролем при определении растворенных липидов - четвертый органический зкстракт из морской воды.
Определение взвешенных и растворенных углеводов проводили триптофановым методом (с L-триптофаном), для которого характерна независимость образующейся окраски анализируемых пробот величины солености в широком диапазоне изменения этого показателя (0- 35%o). Если в пробах морской воды содержание нитратов превышало 10 мкМ, то делали поправкуна изменение интенсивности окрашивания Eberlein, Hammer, 1980).
Определение растворенных и взвешенных НК и нуклеотидов основано на расщеплении олиго- и полинуклеотидов при кислотном гидролизе в 0,5N HClO, на мононуклеотиды, которые интенсивно поглощают свет в ультрафиолетовой
области спектра при 260 - 280 нм.
Результаты
BLOXMLeCKLÈ COCraEPOB. B. Ta5.1H на рис.2 представлено распределение концентраций биохимических компонентов POB и их относительного содержания (в %) в различных районах Охотского моря. Концентрации РОВ установлены, исходя из данных по распределеHC Copr. в рассматриваемом раионе Агатова и др., 1996).
Таблица П. Содержание биохимических компоиентов POB (минимальные и максимальные значения) в Охотском море в летний период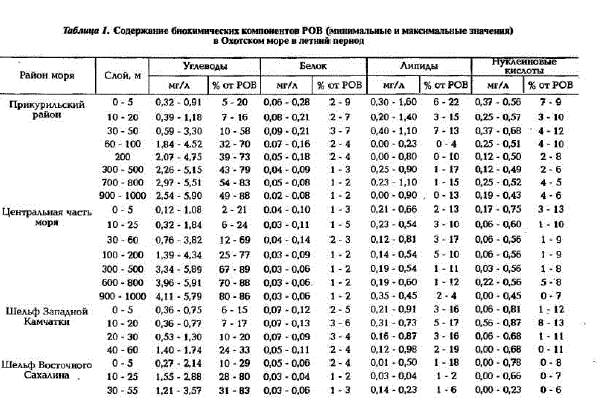
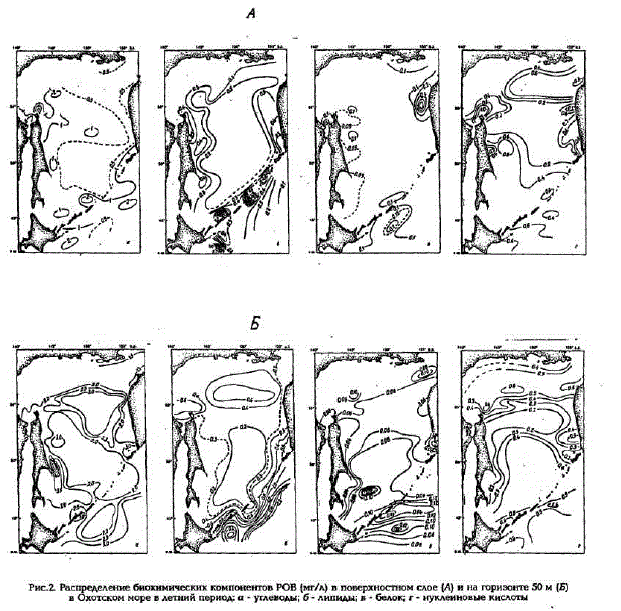 Рис.2. Распределение биохимических компонентов POB (мг/л) в поверхностном слое (А) и на горизонте 50 м (Б) в Охотском море в летний период о - углеводы; б - липиды; в - белок г-нуклеиновые кислоты
Рис.2. Распределение биохимических компонентов POB (мг/л) в поверхностном слое (А) и на горизонте 50 м (Б) в Охотском море в летний период о - углеводы; б - липиды; в - белок г-нуклеиновые кислоты
В верхнем деятельном слое (до 200 м) сумма углеводов, белков, липидов и НК в среднем не превышает 60% от POB (см.табл.1).
Как правило, основным компонентом POB являются углеводы, причем их количество увеличивается с глубиной, особенно это ярко выражено в Прикурильском районе и центральной части моря, где концентрации их возрастают от поверхности до глубины 1000 м в 5 раз. Благодаря этому, в глубинных водах сумма основных биохимических компонентов составляет 90 - 95% от РОВ (см.табл.1). В фотическом слое концентрации углеводов находятся в пределах 0.3 - 1,3 мг/л, исключение составляют воды Сахалинского шельфа, где их содержание увеличивается до 3,0 мг/л; минимальные же концентрации характерны для вод Камчатского шельфа. В мелководных районах (до 30 м) вертикальное распределение углеводов довольно равномерно, кaк правило, с небольшим увеличением их концентраций кодну (рис.3,a). Тогда как в Прикурильском районе и в центральной части моря распределение их по вертикали при общей тенденции увеличения концентраций с глубиной очень неравномерно, с промежуточными максимумами и минимумами (см.рис.3, б).
Межгодовая изменчивость концентраций углеводов повсему морю незначительна, она в основном проявляется на мелководных станциях Сахалинского шельфа. Например, концентрации углеводов здесь в 1994 г. были в 2 - 2,5 раза выше, чем в 1992 г.
Наряду с углеводами большой вклад в POB (до 20%) вносят липиды (см.табл.1, рис.2.б). В отличие от углеводов максимальные их концентрации (выше 1 мг/л) характерны для фотического слоя вод Прикурильского района, а минимальные (менее 0,03 мг/л) - для вод Сахалинского шельфа. Вертикальное распределение липидов на мелководье Камчатского и Сахалинского шельфов довольно равномерно (рис.3,a), тогда как в глубоководной части моря на вертикальном профиле
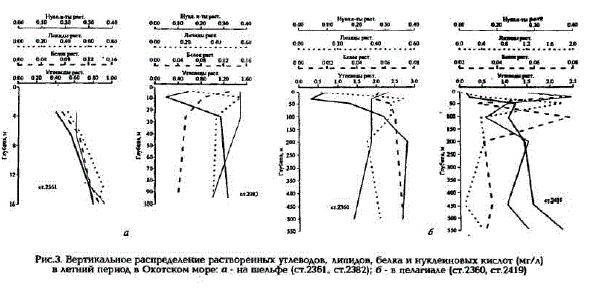
Рис.3. Вертикальное распределение растворенных углеводов, липидов, белка и нуклеиновых кислот (мг/л) в летний период в Охотском море: а - на шельфе (ст.2361, ст.2382) б - в пелагиале (ст.2360, ст.2419)
наблюдаются промежуточные максимумы и минимумы их концентраций при общей тенденции уменьшения с глубиной (см. рис.З,б).
Межгодовая изменчивость концентраций растворенных липидов довольна значительна. Например, в Прикурильском районе и на Сахалинском шельфе концентрации липидов в 1994 г. были в 2 - 6 раз меньше, чем в 1993 г. А в районе банки Кашеварова (ст.2223) в 1994 г. их концентрации, по сравнению с 1993 г., когда были зафиксированы максимальные величины (0.690 мг/л), понизились на порядок.
Доля белков в POB минимальна - в среднем 4 - 7% (см.табл.1). Наибольшие их концентрации характерны для фотического слоя вод в Прикурильском районе (до 0,28 мг/л), а наименьшие (около 0,03 мг/л) - для глубинных вод пелагиали и Сахалинского шельфа (см. рис.2,в). Вертикальное распределение белка на шельфе Камчатки и Сахалина довольно равномерно и концентрации его практически не изменяются от поверхности до дна (см.рис.3,п). В пелагиали же, как правило, содержание белка увеличивается в слое скачка плотности и на границе между тешлым (ТПС) и холодным промежуточными слоями (ХПС). Концентрации белка в глубинных водах в 2 - 3 раза меньше, чем в фотическом слое (см. рис.3.б).
Межгодовая изменчивость концентраций растворенного белка и относительного содержания его в РОВ Невелика.
Доля нуклеиновых кислоп в POB вод Охотского моря значительно больше (до 13%), чем вдруTHx MopsHx AraToBa, TopryHo Ba, 1991; AraToBa H. Ap., 1994; Karl, Bailiff, 1989). Mix KoHleHTpaTMIK "Haсто в несколько раз превышают содержание белка в POB (см.табл.1). Максимальные концентрации НК (до 0,8 мг/л) характерны для вод Камчатского шельфа и Прикурильского района. Наиболее богат HK фотический слой вод шельфа и пелагиали, ниже которого концентрации их равномерно уменьшаются с глубиной, оставаясь довольно высокими (см. рис.3). Однако в слое кислородного минимума на глубинах 600-800 м зафиксировано увеличение концентраций НК по сравнению с выше- и нижележащими слоями. Необходимо отметить очень высокую межгодовую изменчивость концентраций НК, особенно на шельфах Камчатки и Сахалина. В 1992 г. даже в фотическом слое концентрации НК были минимальными, вплоть до аналитического нуля; в 1994 г., напротив, - максимальными; в 1993 г. наблюдались промежуточные значения концентраций НК.
Биохимический состав ВОВ. Данные о распределении основных биохимических компонентов ВОВ в различных районах Охотского моря представлены в табл.2. и на рис.4. Концентрации ВОВ определены, исходя из распределения концентраций взвешенного Copr Aratoba M.Ap, 1996). Borличие от POB, сумма вышеуказанных взвешенных биохимических компонентов практически равна суммарному содержанию ВОВ (до 90 - 95%),
Основным биохимическим компонентом ВОВ является белок, доля которого в ВОВ может составлять более 60% (см.табл.2). Максимальные концентрации белка характерны для фотического слоя вод как шельфа, так и пелагиали. Наибольшие величины (около 0,300 мг/л) отмечены в районах Курильской гряды и Сахалинского шельфа (см. рис.4,в). Для вертикального распределения взвешенного белка во всех районах моря характерно резкое уменьшение концентраций в верхнем слое (0-50 м) и довольно равномерное их распределение в более глубоких слоях (рис.5). Изменения относительного содержания белка с глубиной незначительны, причем в промежуточных водах на некоторых горизонтах оно даже может превышать таковое в фотическом слое (см.табл.2),
Межгодовая изменчивость концентраций взвешенного белка незначительна и наиболее ярко выражена на мелководье Камчатского и Сахалинского шельфов, а также в фотическом слое
Таблица 2. Содержанне биохимических компонентов вов (минимальные и максимальиые значения) в Охотском море в летний период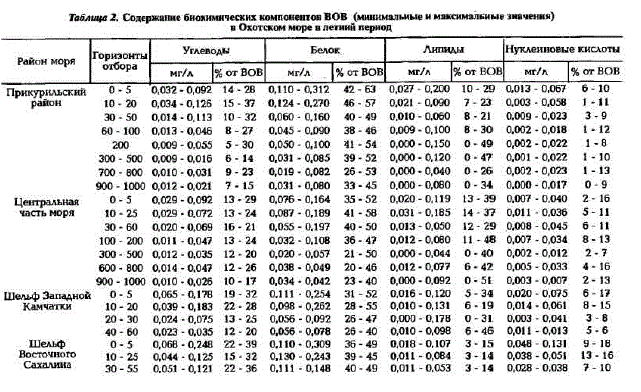
Доля углеводов в ВОВ также весьма значительна (см.табл.2), особенно на мелководье и в фотическом слое пелагиали. Максимальные концентрации углеводов (до 0,240 мг/л) зафиксированы на Сахалинском шельфе, а минимальные (около 0,010 мг/л) - на глубинах более 200 м в Прикурильском районе и в центральной части моря (см.рис.4,a). При общей тенденции уменьшения концентраций углеводов с глубиной распределение их по вертикали очень неравномерно, с промежуточными максимумами в слое 600 - 800 м и минимумами в слое 300 - 500 м (см. рис.5). Следует отметить, что только в водах Прикурильского района происходит уменьшение с глубиной и относительного содержания взвешенных углеводов (см. табл.2), в остальных же районах этот показатель практически не меняется во всей толще вод.
Межгодовая изменчивость концентраций углеводов хорошо выражена только в фотическом слое, максимальные их величины наблюдались летом 1993 г. и были в процентном отношении сопостовимы с содержанием взвешенного белка. В 1992 и 1994 гг. концентрации взвешенных углеводов в 2 раза превышали содержание белка в ВОВ.
Сопоставимый с углеводами вклад в ВОВ вносят и липиды (см.табл.2). Максимальные концентрации липидов зафиксированы в фотическом слое, при этом следует отметить очень большой разброс их значений в пределах исследуемой акватории (см. рис.4,б). Этот разброс отмечался в каждой из трех выполненных в разные годы съемок. Максимальные концентрации взвешенных липидов, так же как и растворенных, наблюдались в фотическом слоевод Прикурильского района и Камчатского шельфа (см.табл.2). Минимальные же значения концентраций взвешенных липидов, равно как и их относительного содержания, отмечались в водах Сахалинского шельфа (см. рис.4,б). Большая неоднородность проявляется и в их вертикальном распределении (cM.pMC,5).
При отмеченном выше "пятнистом" пространственном распределении липидов практически во всех слоях трудно судить о межгодовой изменчивости их содержания.
Доля нуклеиновых кислолп в ВОВ минимальна - в среднем менее 10% . Наибольшие концентрации НК, также как и других биохимических компонентов, отмечены в фотическом слоевод мелководья и пелагиали. Максимальные величины (около 0,130 мг/л) зафиксированы на мелководье Сахалинского шельфа (см. рис.4. г). Вертикальное распределение концентраций НК в пелагиали подобно таковому для взвешенных белков и углеводов (см.рис.5). На Камчатском и Сахалинском шельфах происходит резкое уменьшение концентраций НК от поверхности ко дну (см. рис.5,a).
Труднодать оценку межгодовой изменчивости для НК, также как и для липидов, из-за большой пространственной неоднородности их распределения.
Показателем трансформации ОВ в морских экосистемах является не только колебание биохимического состава РОВ и ВОВ, но и изменение соотношения между растворенными и взвешенными формами соответствующих биохимических компонентов. Например, на шельфе и в открытой части моря на взвешенные углеводы в фотическом слое приходится 10 - 30% от растворенных углеводов, а в глубинных водах - всего 0,5 - 2% от содержания последних. Особенно интересно соотношение взвешенного и растворенного белка. Как правило, на шельфе и в фотическом слое пела
Рис.4. Распределение биохимических компонентов ВОВ (мг/л) в поверхностном слое (А) и на горизонте 50 м (Б) в Охотском море в летний период о - углеводы; б - липиды в - белок г-нуклеиновые кислоты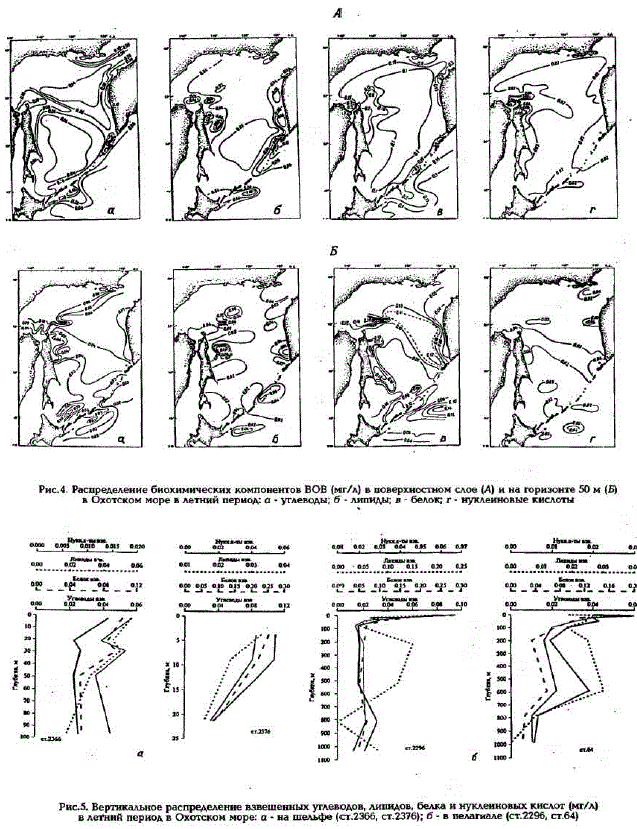 Рис.5. Вертикальное распределение взвешенных углеводов, липидов, белка и нуклеиновых кислот (мг/л) в летний период в Охотском море: а - на шельфе (ст.2366, ст.2376); б - в пелагиале (ст.2296, ст.64)
Рис.5. Вертикальное распределение взвешенных углеводов, липидов, белка и нуклеиновых кислот (мг/л) в летний период в Охотском море: а - на шельфе (ст.2366, ст.2376); б - в пелагиале (ст.2296, ст.64)
гиали (см. табл.1, 2) содержание взвешенного белка в 1,5-2 раза выше, чем растворенного. С глубиной в некоторых районах моря доля взвешенного белка падает до 30-50% от содержания растворенного белка. В водах Сахалинского шельфа концентрации взвешенного белка в 2 - 5 раз больше, чем растворенного, тогда как в водах Камчатского шельфа эти соотношения сохраняются только до глубины 20 м, а ниже концентрации растворенного белка возрастают относительно содержания взвешенного.
Следует отметить, что вышеприведенные соотношения не карактерны для раионов со значительной изменчивостью гидрологических условий - Сахалинского залива и проливов Курильской
HNI. Pಣ್ಣ Сахалинском заливе четко выявляется влияние притока пресных вод р. Амур на распределение растворенных и взвешенных компонентов OB. Поступление речных вод способствовало увеличению концентраций всех биохимических веществ. Максимальные концентрации растворенных компонентов: углеводы - 1,26 мг/л, белки- 0,213 мг/л, липиды - 0,358 мг/л, НК-1,86 мг/л; взвешенных углеводы - 0,201 мг/л, белки - 0,397 мг/л, липиды - 0,105 мг/л. НК - 0210 мг/л-
Неустойчивый гидродинамический режим в проливах Курильской гряды обуславливает не только большую межгодовую изменчивость концентраций всех биохимических компонентов ОВ, но и приводит к нарушению описанных выше особенностей вертикального их распределения в водах Прикурильского района.
В табл.3 представлены данные по распределению биохимических компонентов РОВ и ВОВ в поверхностном микрослое (ПМС). Концентрации растворенных биохимических веществ в ПМС в 1,5 - 6 раз больше, чем в подстилающем его слое (см.табл.3), тогда как концентрации взвешенных биохимических компонентов превышают их содержание в подстилающем слое в 2 - 12 раз (см.табл.3). Следует отметить, что на некоторых станциях концентрации растворенных углеводов, липидов (ст.2292) и белка (ст.2378) в подстилающем слое были выше, чем в ПМС, тогда как концентрации всех взвешенных биохимических компонентов всегда были значительно выше в TMO.
Таблица 3. Коицеитрацин биохимических компонентов РОВ и ВОВ (мгіл) в повершостном микрослое (в числителе) и слое 00-0,5 м (в знамеинтеле) в Охотском море
No стан- Углеводы Белок Липиды Hಞ್ಞಟಿ Углеводы Белок Липиды Hಳ್ದುಟಿ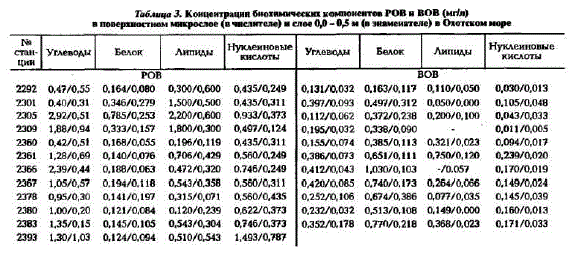
Обсуждение результатов исследования
Изменение концентраций растворенных и взвешенных биохимических компонентов ОВ должно быть тесно связано с колебанием биологических показателей морской KEMIEI
В клетках фитопланктона (в пересчете на сухой вес) 50% приходится на белок, 30% - на углеводы, 10% - на липиды и 10% - на НК, а в клетках бактериопланктона: 50% - на белок, 12% - на углеводы, 8% - на липиды и 30% - на НК (Парсонс и др., 1982), поэтому интенсивность процессов первичного продуцирования B OCHOBHOM AO MYKHöl отражаться LLLLLL LLLLLLL LLL LLL LLGLLL aLLLLLSLLLLLLLS ных углеводов. Действительно, в тех районах, где наблюдали максимальные величины фитопигментов, первичной продукции и биомассы фитопланктона (Микаэлян и др., 1995; Налетова и др., 1996) в фотическом слое, были зарегистрированы и максимальные концентрации не только взвешенных, но и растворенных углеводов. При этом доля углеводов в ВОВ могла достигать 50%.
Изменчивость содержания углеводов зависит также от интенсивности деструкционных процессов. С этой точки зрения интересны данные по вертикальному распределению концентраций растворенных углеводов в открытой части моря (см. рис.3.б), где были зафиксированы интенсивные продукционные процессы. При общей тенденции увеличения их концентрации с глубиной, в слое 10 - 40 м происходит уменышение количества растворенных углеводов до величин в 1,5 - 2 раза меньших, чем на поверхности или в слое максимальной продукции. Это уменьшение связано с потреблением растворенных углеводов микрогетеротрофами, так как на этих же горизонтах повышаются концентрация взвешенного белка и скорость потребления кислорода микропланктоном (Агатова, Лапина, 1996). Ранее для вод Северной Атлантики было показано, что в слое максимальных концентраций бактериопланктона содержание растворенных углеводов минимально
(Burney et al., 1979). Такой промежуточный минимум мы наблюдали также в районе материкового склона и глубоководной части Берингова моря Агатова и др. 1995, в пелагиали Черного моря Агатова и др., 1989, Агатова, Торгунова, 1991). Ханда (Handa, 1970) также указывал на более быстрое потребление и разложение углеводов в фотическом слое.
Интенсивность продукционно-деструкционных процессов влияет и на соотношение между растворенными и взвешенными формами углеводов. Так, в местах активного фотосинтеза и большой биомассы микрофитопланктона взвешенные углеводы составляют 10- 30% от растворенных. В фотическом слое, где скорость деструкции превышает скорость первичного продуцирования, а также в глубинных водах взвешенные углеводы от растворенных составляют всего 0,5 - 2%. В глубинных водах такой низкий процент взвешенРис.б. Распределение растворенных углеводов (мг/л) на раз- ных углеводов относительно растворенных резе от сахалинского шельфа (ст.7 около п-ова терпения) к связан с тем, что здесь значительно возраста
проливу Фриза с выходом в тихоокеанскую частъ Прику- ет концентрация последних.
рильского района (ст.бВ) Увеличение растворенных углеводов с глубиной в 1,5-2 раза было отмечено вводах Индийского океана в 1963 г. (Handa, 1966) и в 3 раза - в водах Берингова моря в 1985 г. (Sakugawa, Handa, 1985). Однако концентрации растворенных углеводов в глубинных водах, согласно цитируемым выше авторам, оказались в 20-50 раз меньше полученных нами концентраций растворенных углеводов в Охотском море. Напримере формальдегиданами было показано, что альдегиды влияют на интенсивность окраски при определении углеводов с L-триптофаном. Альдегиды в морской воде могут полимеризоваться до углеводоподобных продуктов (Schwartz, 1981). Искодя из этого можно предположить, что полученные нами высокие концентрации растворенных утлеводов в глубинных водах связаны с поступлением сюда окисленных нефтепродуктов с шельфа. На разрезе от Сахалинского шельфа к проливу Фриза с выходом в тихоокеанские воды (рис.6) по распределению растворенных углеводов хорошо прослеживается такое распространение нефтепродуктов. Большая межгодовая изменчивость содержания углеводов, как растворенных так и взвешенных, отмеченная на Сахалинском шельфе, главным образом связана с интенсивностью нефтеразработок здесь. При проведении буровых работ происходит увеличение концентрации взвешенных частиц на которых сорбируются растворенные органические вещества, в частности высокополимерные полисахариды, что приводит к уменьшению концентрации углеводов в растворе и увеличению во взвеси.
При интенсивных процессах первичного продуцирования соответственно в фотическом слое происходит увеличение содержания взвешенного белка, причем не только за счет роста численности и биомассы фитопланктона, но и также за счет увеличения численности и биомассы бактериопланктона (Сорокин и др., наст.сб.). Повышение численности бактериопланктона в результате интенсивного первичного продуцирования сказывается на увеличении количества не только взвешенного белка, но и растворенного. Последнее скорее всего связано с большим вкладом нано- и пикоформ в бактериопланктон. Эти формы свободно проходят через фильтры CF/Fимогут быть обнаружены только с помощью седиментационных ловушек [Karl et al.,1988; Коike et al., 1990). С другой стороны, повышение содержания растворенного белка может происходить за счет продуктов метаболизма фотосинтезирующего планктона, так как в холодных водах фитопланктон преимущественно синтезирует белок, а синтеза липидов и сахаров практически не проMcxoAMT [Li, Harrison, 1982].
Уменьшение по вертикали доли взвешенного белка относительно растворенного связано с изменением соотношения микро- и пикоформ планктона вследствие уменьшения биомассы микропланктона с глубиной (Сорокин и др., 1996). Ранее такая же закономерность была показана для глубоководной части Черного моря (Агатова, Торгунова, 1991). Поэтому можно предположить, что понижение с глубиной концентрации растворенного белка связано соответственно с уменьшением биомассы пикоформ. Однако наличие промежуточных максимумов растворенного и взвешенного белка указывает на то, что в скачке плотности и на границе между ХПС и ТПС происходит скопление бактериопланктона. Ранее (Агатова и др., 1995) мы наблюдали такого родаскопления в водах Берингова моря - в слоях скачка плотности и минимума кислорода. Для вод Северной Атлантики еще в 1976 г. (Karlet al., 1976) по увеличению концентраций АТФ было также обнаружено скопление бактериопланктона в слое минимума кислорода.
Увеличение концентраций растворенных и взвешенных нуклеиновых кислота и их доли относительно белка в этих же слоях не только подтверждает скопление бактериопланктона, но и позволяет предположить его активное размножение, поскольку при размножении бактериальных клеток значительно возрастает доля НК в РОВ и ВОВ (Karl, 1978).
В фотическом слое при интенсивном первичном продуцировании зафиксированы максимальные концентрации НК. Однако очень большой процент НК в ОВ и особенно низкое соотношение белок/НК могут быть только в половых продуктах. Наличие высоких концентраций НК в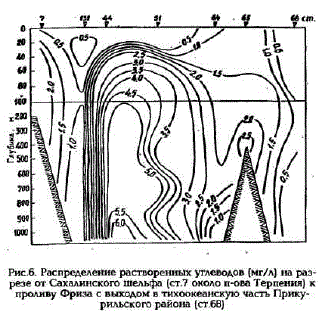
POB мы связываем с местами интенсивного нереста гидробионтов, так как во всех известных рыбопродуктивных районах Охотского моря (Шунтов, 1995) были зафиксированы наибольшие концентрации и растворенных и взвешенных НК. Поэтому именно для НК отмечена самая большая по сравнению с другими биохимическими компонентами межгодовая изменчивость.
Довольно значительные межгодовые колебания характерны и для растворенных липидов, особенно в рыбопродуктивных районах Камчатского шельфа и Прикурильского района. Интересно, что при этом из года в год вдоль Курильской гряды - от Кожных Курил к Северным - отмечается уменьшение концентраций липидов доминимальных (0,1 мг/л). Эти данные находятся в прямой зависимости от распределения здесь скоплений рыбы (Баланов, Радченко, 1995) и зоопланктона (Мусаева, Колосова, 1995). Тесная связь липидов с биомассой зоопланктона, по-видимому, обуславливает и большую пятнистость в пространственном распределении взвешенных липидов.
Неравномерное распределение взвешенных и растворенных липидов по вертикали, также как и других биохимических компонентов (см. рис.4), зависит от изменения биологических факторов. Так, промежуточные максимумы концентраций липидов зафиксированы на тех же горизонтах, на которых обнаружены максимальные концентрации белка, углеводов и НК, т.е. в местах скопления микро- и пикопланктона. Картина такая же, которую мы наблюдали ранее в пелагиали Черного и Берингова морей (Агатова, Торгунова, 1991; Агатова и др.,1995). В водах северной части Тихого океана и Черного моря было обнаружено скопление ВОВ в слоях на глубине около 500 м, с которым совпадали повышенные концентрации взвешенных липидов, выделенных из обитающих там микрогетеротрофов (Wakeham, 1995).
Таким образом, изменение концентраций растворенных и взвешенных биохимических компонентов и их соотношений в различных районах Охотского моря определяется колебанием биологических показателей экосистемы. Влияние же гидрологических условий на изменчивость биохимического состава РОВ и ВОВ достаточно очевидно только в Сахалинском заливе, на банке Кашеварова и в проливах Курильской гряды.
Так, распространение пресных вод р. Амура в Сахалинском заливе привело к увеличению концентраций ВОВ и всех вышеперечисленных его биохимических компонентов в поверхностном слое, а концентраций РОВ и его компонентов - в придонном слое. Это объясняется созданием биологического барьера в зоне смешения речных и морских вод за счет увеличения биомассыгетеротрофов и их активности (Агатова, Лапина, 1996). Аналогичную картину мы наблюдали в Анадырском заливе Берингова моря (Агатова, Лапина, 1994).
В свою очередь, активное перемешивание вод в проливах Курильской гряды и над банкой Кашеварова обуславливает однородный характер вертикального распределения биохимических компонентов. На банке Кашеварова концентрации и растворенных и взвешенных веществ практически не изменяются от поверхности до дна. В проливах Курильской гряды только концентрации биохимических компонентов ВОВ в фотическом слое до некоторой степени зависят от интенсивности биологических процессов. Ниже этого слоя концентрации и взвешенных и растворенных биохимических компонентов практически не меняются с глубиной, за исключением растворенных углеводов. Однако увеличение их концентраций с глубиной незначительно по сравнению с таковой в мористой части Прикурильского района.
Отмеченное резкое увеличение концентраций РОВ и ВОВ (Агатова и др., наст. сб.) и их биохимических компонентов в ПМС по сравнению с подстилающим слоем обусловлено, вероятно, физико-химическими свойствами этого слоя, способного удерживать полярные молекулы [Hunter, Liss, 1981], что, в свою очередь, благоприятствует концентрированию в нем микропланктона. Аналогичные концентрации биохимических компонентов в ПМС наблюдались ранее в водах Берингова моря (Агатова и др., 1995) и в районе тихоокеанского побережья КалифорнийскоTo IIowyOCTpoBa Henrichs, Williams, 1985.
Выводы
1. Изменения концентраций основных биохимических компонентов РОВ и ВОВ, а также их соотношений главным образом определяются биологической активностью автотрофных и гетеротрофных обитателей экосистем шельфа и глубоководных районов Охотского моря.
2. Основными биохимическими компонентами РОВ и ВОВ в Охотском море являются углеводы и белок соответственно. При интенсивном первичном продуцировании доля углеводов в ВОВ возрастает и становится соизмеримой с таковой белка.
3. Высокое содержание растворенных и взвешенных НК и превышение их содержания над концентрациями растворенного белка, а также высокие концентрации растворенных липидов в районе Южных Курил и на шельфе Западной Камчатки находятся в тесной связи с наличием здесь скоплений рыбы и зоопланктона.
4. Характер горизонтального и вертикального распределения основных биохимических компонентов РОВ и ВОВ нашельфе и в пелагиали Охотского моря обусловлен главным образом биологическими факторами. Влияние же гидрологических условий на эти показатели довольно сильно выражено в Сахалинском заливе, проливах Курильской гряды и над банкой Кашеварова.
5. Концентрации основных биохимических компонентов РОВ и ВОВ в поверхностном микрослое значительно больше, чем в подстилающем его слое, что связано с физико-химическими CB01CTBaMH TIMC,
Работа выполнена при финансовой поддержке РФФИ (код проекта 96-05-64576).
A.M.Aгатова, H.M.Павлова, H.M.Topгушева
Смотрите также






