Экология молоди горбуши в эстуарно-прибрежный период жизни
Молодь, скатывающаяся из рек, имеющих лиманы, задерживается в них для нагула и осморегуляторной адаптации. Пребывание в эстуариях молоди горбуши длится от 3 недель до 2—3 мес. Было исследовано 55 окз. в июне-июле. За это время молодь в среднем достигает 45,7 (+ 1,3) мм и 0,82 (+ 0,07) ги происходит ее адаптация к воде сразличной соленостью и физиологическая перестройка организма.
Период перехода из пресных вод в морские для всех видов лососей, в том числе и для горбуши, является наиболее важным в формировании численности поколений. Исследования, проведенные E.E. Изергиной с соавторами (2009) в периодадаптации и смолтификации горбуши в лиманер. Ола, показали, что в организме рыб происходят серьезные физиологические трансформации. Для оценки адаптивных возможностей молоди горбушик изменяющимся условиям среды авторы исследовали изменения, происходящие в качественном составе форменных элементов крови у «дикой» молоди горбушив сстественных условиях ската при переходе из пресных вод в морские. В целях поддержания гомеостаза кровь отвечает изменениями своего состава на любые экзогенные и эндогенные воздействия. Изучение морфологической структуры периферической крови молоди горбуши из участков эстуария сразличной соленостью позволило характеризовать динамику развития осмотического гомеостаза в период ее смолтификации в эстуарный и ранний морской периоды жизни.
Установлено, что уровень эритропоэза в процессе адаптации горбушик воде повышенной солености чрезвычайно высок — происходит интенсивнос образование новых эритроцитов различными способами: как в органах кроветворения, так и путем прямого деления эритроцитов, в том числе отмечено наличие безъядерных клеток, участвующих в транспорте кислорода. То есть в процессе смолтификации происходило разрушение зрелых эритроцитов и их замена молодыми формами, а уровень эритропоэза в процессе адаптации к соленой воде был чрезвычайно высок (Изергина и др., 2009).
При переходе в соленую воду количество бластных клеток эритроидного ряда увеличивается вдвое. Общее количество лейкоцитов у молоди горбуши увеличивается в период tala Tai Li K. условиям IIOBEILIIEHHOH COJIEHOCTH, HTO является адекватной реакцией иммунной системы на смену среды обитания. Доля лимфоцитов снижается до 60% и более, увеличивается доля промиелоцитов и сегментоядерных клеток. Количество тромбоцитов у молоди горбуши при переходе из пресной воды в соленую составляет более 20%, снижаясь по мере роста и завершения процесса смолтификации до 2%.
Молодь горбуши в начальный период смолтификации более резистентна к условиям резко увеличивающейся солености воды, чем молодь кеты. Это, вс
роятно, обусловлено особенностью приспособительных реакций горбуши, молодь которой в момент перехода в соленую воду характеризуется пассивным скатом и не совершает активных перемещений из выносящего потока в зоны переменной солености, которые предпочитает молодь кеты (Изергина, Изергин, 2009). То есть пассивный вынос мальков горбуши в воду морской солености без адаптации в воде эстуариев, очевидно, не вызывает ее массовой гибели. В большей степени это свойственно скатывающейся молодикеты.
После выхода в прибрежье мальки горбуши распределяются в приливноотливной зоне. Период пребывания молоди в прибрежье может составлятъ от нескольких суток до 80—90 дней, причем откочевка горбуши из приливно-отливной зоны происходит несколько раньше, чем кеты. В течение июня и до середины июля молодь обитает в прибрежных, хорошо прогреваемых бухтахизаливах. В Тауйской губе это бух. Мелководная, Гертнера, Нерпичья, Токарева, расположенные в зал. Одян и Мотыклейский. В зал. Шелихова в июне молодь держится в прибрежнойзоне шириной до 4 миль и образуетскопления в небольших бухточках и заливах. Температура воды в них прогревается до 10—12°С. Откочевка в море начинается с середины июля. Во второй половине июля молодь горбуши
Принято считать, что высокая ледовитость в период выхода молоди горбушив прибрежье негативно влияет на формирование численности ее поколений. Это подтверждается и нашими данными для северо-восточного участка материкового побережья Охотского моря. Например, прямые наблюдения в прибрежье показали, что длительная задержка льдов в приустьевых зонах лососевых нерестовых рек приводит к резкому увеличению смертности горбуши. Площадной учет, проведенный в год высокой ледовитости моря (1995 г.), показал, что во время отливов в прибрежье зал. Мотыклейский на 1 м° грунта насчитывалось до 8 погибших мальков, а коэффициент возврата горбуши этого поколения (1994 г.) был одним из наименьших за весь многолетний период наблюдений — около 0,5%.
Аналогичный пример негативного влияния на выживаемость горбуши юговосточного Сахалина в результате мощного выноса льдов из северной части Охотского моря приведен О.Ф. Гриценко с соавторами (1987. С. 43). Произошло резкое снижение температуры воды (до 0,4°С), отмечалась гибель молоди горбуши от охлаждения и давления льдов в прибрежье. Численность возврата горбуши этого поколения была низкой.
Ледовитость моря является основным фактором, определяющим выживаемость поколений североохотоморской горбуши. Массовый выходгорбушив прибрежье приходится на первую половину июня. В том случае, если в Тауйской губельды сохранялись до конца IIдекады июня, выживаемость поколений была низкой. Зависимость выживаемости молоди горбуши от относительной площади льдов в Тауйской губе в первой половине июня хорошо описывается экспоненциальной зависимостью (рис. 49).
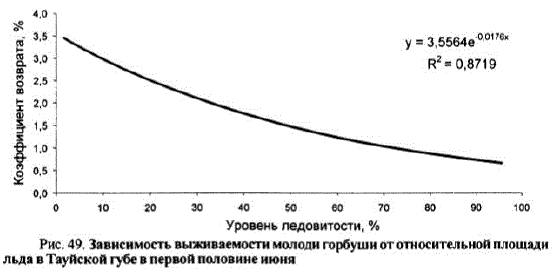
Рис. 49. Зависимость выживаемостимолоди горбуши от относительной площади льда в Тауйской губев первой половине июня
Хорошие коэффициенты выживаемостиугорбушинаблюдаются в тех случаях, когда в период ее массового выхода в прибрежье относительная площадь льдов в Тауйской губе была менее 25%. Напротив, высокая смертность была утех поколений, которые скатывались в Тауйскую губу, заполненную льдами (см. рис. 49).
Однако объяснять изменения уровня выживаемости горбуши только влиянием ледовитости было бы не совсем объективно. Например, коэффициент выживаемости поколений горбуши, мигрировавших в Тауйскую губу, свободную ото льда, варьировал в довольно широких пределах — от 1,7 до 5,6%. Это свидетельствует о наличии и других факторов, определяющих успешность выживания горбушив начальный морской период жизни.
Вероятно, не последнюю роль в выживании молоди горбуши в прибрежье играет ее обеспеченность пищей. В качестве косвенного индикатора состояния кормовой базы молоди горбушинами был привлечен такой показатель, как концентрация хлорофилла-«а» в прибрежных водах. Выполненные исследования показалитесную связь между концентрацией хлорофилла-«а» в водах Тауйской губы во второй половине мая и выживаемостью молоди горбуши (рис. 50),
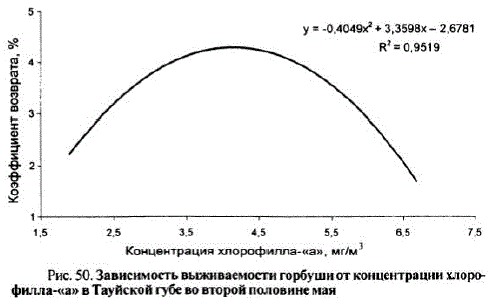
Рис. 50. Зависимость выживаемости горбуши от концентрации хлорофилла-«а» в Тауйской губево второй половинемая
150
В этом случае, видимо, проявляется инерционная зависимость — оптимальная концентрация фитопланктона в конце мая обеспечивает формирование достаточной кормовой базы для молоди в июне. Хорошая выживаемость молоди горбуши наблюдалась при концентрациях хлорофилла-«а» от 2,5 до 5,5 мг/м". Другими словами, в том случае, если концентрация хлорофилла-«а» составляла менее 2 мг/м", уровень развития фито- и зоопланктона, очевидно, не может обеспечить в полной мере пищевую потребность молоди доступным кормом, определяющую выживаемость молоди горбуши. Напротив, когда содержание хлорофилла-«а» составляет 4—5 мг/м", мы имеем дело со вспышкой фитопланктона, за которой через некоторое время последуетрост биомассы зоопланктона. Кроме того, не исключаем негативное воздействие вспышек фитопланктона на дыхательную функцию молоди, приводящую, вероятно, к увеличению смертности мальков,
По нашему мнению, исследование только океанической стороны жизни лососей — это односторонний подход к пониманию причин, лежащих в основе формирования их численности. Конечно, существуют весомые аргументы в виде огромного массива литературных данных, довольно полно характеризующих пресноводный и ранний морской периоды жизни молодилососей как на Азиатском, так и на Североамериканском побережьях Тихого океана. Тем не менее мониторинговые исследования эстуарно-прибрежного периода жизнилососей необходимо продолжать, и это позволит нам держать под контролем ситуацию с выживаемостью молодилососей в наиболее критический периодонтогенеза. Приведенные частные примеры установленных нами связей свидетельствуют о важности долгосрочных мониторинговых исследований пресноводного и эстуарно-прибрежного периодов жизнилососей, создающих базу для моделирования их динамики численности.
Таким образом, углубленные комплексные исследования эстуарных зон лососевых рек, включающие изучение совокупности гидрологических и гидрохимических факторов, выявление причин смертности молодилососей на физиологическом уровне в период смолтификации, позволят с достаточной степенью достоверности оценивать влияние лимитирующих факторов окружающей среды на популяции лососевых.
Несомненно, что рассмотренные нами показатели не раскрывают всей многогранности вопроса выживаемости горбуши в ранний морской период жизни. Тем не менее, расширение спектра исследований и привлечение для оценки выживаемости новых показателей позволит получать более полные данные и использовать их в прогностических целях.
Тихоокеанских лососей, как виды проходные и далеко мигрирующие, необходимо изучать на всех стадиях жизненного цикла. Реализация жизненной стратегии у них осуществляется в 3 этапа: пресноводный, эстуарно-прибрежный и морской, включая океанический. По эффективности воспроизводства, выживания на пресноводном этапе жизни и при переходе в морскую средумы получаем оценку численности формирующихся поколений. Исследование факторов, определяющих их конечную численность в морской период жизни, дает нам возможность осуществлять управление ресурсами и рационально их эксплуатировать.
Смотрите также






