Внутривидовая структура горбуша
Географическая подразделенность. Как вид горбуша образует устойчивые региональные комплексы: Западной и Восточной Камчатки, Сахалина, Южных Курильских островов, материкового побережья Охотского моря, Амура, Приморья, характеризующиеся собственной динамикой численности, биологическими показателями и сроками нерестового хода.
Горбуша — единственный вид из тихоокеанских лососей, имеющий две практически неперекрывающиеся линии генераций — репродуктивно обособленные во времени смежные поколения четных и нечетных лет. Поколения четных и нечетных лет не смешиваются в природе, и их можно рассматривать как темпоральные группировки вида. Как правило, в разных регионах доминирует какая-нибудь одна из линий, чаще более многочисленны поколения нечетных лет. В принципе, разные линии поколений горбуши можно рассматривать как виды-двойники, поскольку они отвечают критериям, предложенным для дифференциации животных этих категорий: репродуктивная изоляция, сходная морфология, симпатрия (Майр, 1971, 1974 Грант, 1980, Степанян, 1983, Иванков, 1997). Линии горбуши четных и нечетных лет имеют устойчивые и достоверные отличия по экологии, морфологии, кариотипам, маркерам генов (Двинин, 1952, Смирнов А. И., 1975, Картавцев и др., 1981, Салменкова и др., 1981, Черешнев, 1981; Алтухов и др., 1983, 1987, 1997; Горшков, 1979; Горшкова, Горшков, 1983; Ефанов, 1989; Омельченко, Вялова, 1990; Варнавская,2006;Aspinwall, 1974).
Кроме двух симпатрических репродуктивно изолированных в природелиний горбуша образуетсезонные расы (Иванков, 1967а, 1971; Смирнов А.И., 1975), отличающиеся сроками нерестовой миграции (например, летняя и осенняя), местами размножения, биологическими показателями. Отмечен нерест поздней осенней горбушина выходах грунтовых вод (Канзепарова, 2009), что принципиально отличает ее от горбуши летненерестующей формы и, очевидно, обусловливает морфоэкологическую дифференциацию ее сезонных рас. В СахалиноКурильском регионе установлено наличие трех сезонных группировок: япономорской, летней и осенней охотоморских (Гриценко, 1981). В бас. р. Гижига установлено наличие двух экологических форм горбуши — ранней, совершающей нерестовую миграцию в июне, и поздней, заходящей на нерест в июле. Эти экологические группировки статистически достоверно различаются по биологическим показателям, темпуроста, структуречешуи (Марченко, 2001). Наличие ранней и поздней группировок горбуши установлено и для других рек североохотоморского побережья — Олы и Тауя (Марченко, 1999).
В региональных комплексах обособляются локальные стада (группы популяций), выделение которых основано на различиях динамики численности, комплекса основных биологических показателей, особенностях структуры чешуи, морфооблика и темпов роста. Так, например, по совокупности этих признаков на северном побережье Охотского моря выделены четыре локальных стада: гижигинское, ямское, ольское и тауйское. В указанных стадах североохотоморской горбуши может проявляться клинальная изменчивость по отдельным признакам, например, подлине тела: уменьшение размеров наблюдается в направлении с юго-запада на северо-восток побережья (Марченко, 2004). Кроме того, внутри локальных стад выделяются сезонные или темпоральные группировки, различающиеся сроками подхода. Например, ранняя и поздняя по срокам анадромной миграции горбуша. В свою очередь, сроки подходов могут детерминироваться разными районами нагула в океане: горбуша, нагуливающаяся в более южных районах, обычно подходит на нерестраньше. Для рекрутов горбуши, заходящих из океана на нерест в Охотское море, установлен «волнообразный» характер преднерестовых миграций, обусловливающий подход той или иной региональной группировки в определенный участок охотоморского побережья (Шубин, Коваленко, 2000). В авангарде преднерестовых скоплений, направляющихся в Охотское море, обычно преобладает наиболее ранняя по срокам миграции североохотоморская горбуша. Волновая природа нерестовых миграций горбуши в реки сохраняется в виде дискретных темпоральных группировок, или «ходов»,
33
Следующим уровнем в популяционной организации горбуши являются популяции, приуроченные к отдельным речным бассейнам. Внутри популяций могут выделяться субпопуляции, или демы, тяготеющие к отдельным комплексам нерестилищ — локальным участкам речного бассейна или притокам рек.
Впервые районирование североохотоморского побережья было проведено В.К. Клоковым (1970). В основу его схемы деления были положены географическая обособленность районов, особенности динамики численности, преобладание в уловах отдельных видов и отличия в биологической структуре нерестовых стад лососей. В то же время вышла совместная работа В.К. Клокова и Л.А. Фроленко (1970), посвященная изучению элементарного химического состава чешуи горбуши, подтверждающая предложенную В.К. Клоковым структуру районирования. Согласно этим исследованиям, на северном побережье Охотского моря были выделены три рыбопромысловых района: гижигинский — от р. Авекова до р. Вилига, ямский — отр. Калалагадор. Яма; тауйский — отр. Сиглан до рек Мотыклейского залива.
Дальнейшее уточнение промысловой структурированности региона для горбуши выполнил И.С. Голованов (1983). Основываясь на сравнении основных биологических показателей (длина по Смитту, масса тела и плодовитость) горбуши разных рек, он разделил тауйский рыбопромысловый район на ольский (от р. Сигландор. Ола) и тауйский (от р. Армань до рек Мотыклейского залива) подрайоны с входящими в них группами рек.
Новейшую схему районирования разработали С.Л. Марченко и И.С. Голованов (2001). В ее основу были положены величины ежегодных подходов горбушив реки северного побережья Охотского моря и циркуляция вод деятельного слоя в прибрежной зоне в теплый период (Чернявский, 1981; Чернявский и др., 1981; Шунтов, 2001). Применение факторного анализа позволило классифицировать водоемы североохотоморского побережья и отнести их к нескольким, хорошо обособленным друг от друга, группам рек, совпадающим по своим границам с циркуляциями вод прибрежья (рис. 37). Две из выделенных групп располагаются в Тауйской губе, две — в зал. Шелихова (рис. 38).

Рис. 37. Распределение рек северного побережья Охотского моря подинамике подходов горбуши с 1966 по 2003 г. на плоскости координат. А — Гижигинская группа peK: l — ABeKoBa, 2 — TM»KHTa, 3 — BapxaJIaM, 4 — B6. TapMaHJua, 5 — HasxaH, 6 — VñKaHe, 7 — Студеная, 8 — Таватум, 9 — Широкая, 10 — Пропащая, 11 — Вилига, 12— Калалага, 13 — Кананыга; Б — Ямская группа рек: 14—Туманы, 15 — Наслаган, 16 — Булун, 17 — Угулан, 18 — Тахтояма, 19 — Иреть, 20 — Яма; В — Ольская группа рек: 21 — Сиглан, 22 — Богурчан, 23 – Окурчан, 24— Нюрчан, 25 — Oла; Г—Тауйская группа рек: 26 — Армань, 27 — Ойра, 28 — Яна, 29 —Tayй, 30—Мотыклейка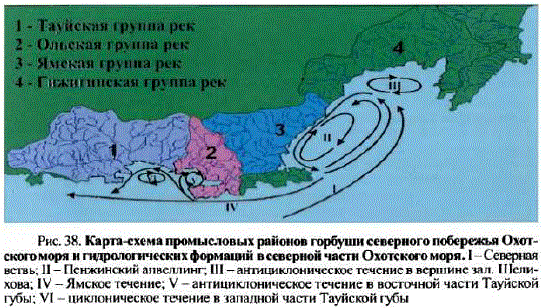
Рис. 38. Карта-схема промысловых районов горбуши северного побережья Охотского моря и гидрологических формаций в северной частн Охотского моря. I—Северная вствь; II— Пенжинский апвеллинг; III— антициклоническое течение в вершинезал, Шелихова; IV— Ямское течение; V — антициклоническое течение в восточной части Тауйской губы; VI— циклоническое течение в западной части Тауйской губы
В прежних границах остались Ольская и Тауйская группы рек. Объем Ямской группы рек был уменьшен за счет перевода двух рек — Калалага и Кананыга — из ее состава в Гижигинскую группу. Из всех выделенных совокупностей наиболее плотную плеяду образуют реки Гижигинской группы. В остальных группах реки располагаются более дисперсно (см. рис. 37).
Расположение плеяд на плоскости координат интерпретируется следующим образом. Ось абсцисс показывает, какая генеративная линия доминирует в той или иной группе рек и отражает вклад доминирующего поколения в общую величину подходов. Ольская и Гижигинская группы рек расположены в противоположных частях декартовых координат. В Ольской группе долгое время (с 1986 по 1998 г.) доминировали генерации четного ряда лет. Вклад доминирующего поколения в подходы горбуши достигал 70,4% от общей величины подходов. В Гижигинской группе на протяжении всего периода наблюдений доминируют поколения ряда нечетных лет. Его вклад в подходы достигает 86,9%. В Тауйской и Ямской группах рек также доминируют поколения ряда нечетных лет, однако уровень подходов горбуши в данные группы рек существенно ниже, чем в реки Гижигинской группы. Кроме того, в отдельные годы в реки Ямской и Тауйской групп наблюдались урожайные подходы в рядучетных лет — 1990 и 1992 г. Поэтому они занимают промежуточное положение между Ольской и Гижигинской группами рек, со смещением в сторону Гижигинской группы. Вклад доминирующего поколения в величину общего подхода в Ямскую группу рек достигал 61,8%, в Тауйскую — 61,2%.
Ось ординат на рис. 37 в общих чертах показывает величину запасов горбуши той или иной реки в группе. Водоемы, расположенные в верхней части группы, обладают минимальными, а в нижней части — максимальными 3ELITECEMH.
Выделение математическим методом промысловых районов поставило нас перед необходимостью проверки реальности их существования. Принимая во внимание тот факт, что в начальный морской период гибнет до 97—99% скатившейся молоди, мы предположили, что именно условия прибрежья являются одним из основных факторов, определяющих конечную величину подхода производителей горбушинатом или ином участке побережья, а в нашем случае — рыбопромысловом районе.
Анализ литературы подтвердил наше предположение: выделенные рыбопромысловые районы по своим границам совпали с гидрологическими формациями вод прибрежья (табл. 42; см. рис. 38).
Таблица 42. Рыбопромысловые районы горбуши и гидрологические формации в прибрежье северного побережья Охотского моря
 Согласно данным В.И. Чернявского (1981), гидрологические формации обладают уникальными физико-химическими характеристиками. Работы, проведенные в Тауйской губе в 1986—1990 гг. (Афанасьев и др., 1991), подтвердили эти данные и, кроме того, показали различия в качественном составе планктонных сообществ западной и восточной частей Тауйской губы. На основании этого можно предположить, что гидрологические образования по своей сути являются отдельными крупными биотопами, а их устойчивость во времении пространстве определяет уровень выживаемости сеголетков и динамику ежегодных подходов горбушина отдельных отрезках побережья.
Согласно данным В.И. Чернявского (1981), гидрологические формации обладают уникальными физико-химическими характеристиками. Работы, проведенные в Тауйской губе в 1986—1990 гг. (Афанасьев и др., 1991), подтвердили эти данные и, кроме того, показали различия в качественном составе планктонных сообществ западной и восточной частей Тауйской губы. На основании этого можно предположить, что гидрологические образования по своей сути являются отдельными крупными биотопами, а их устойчивость во времении пространстве определяет уровень выживаемости сеголетков и динамику ежегодных подходов горбушина отдельных отрезках побережья.
Циклонические водовороты, или апвеллинги, характеризуются мощным течением. Эти циркуляции, за счет создания зоны разрежения в поверхностном слое, способствуют подъему холодных, обогащенных биогенами придонных вод. С одной стороны, подьем вод приводит к обогащению биогенными веществами деятельного слоя, но, с другой — существенно его охлаждает. В период весеннего льдоразрушения квазистационарная система Пенжинского апвеллинга удерживает ледовые поля, вовлекая их в водоворот. Задержка JTEITa вблизи берегов отрицательно сказывается на прогреве поверхностных вод прилегающего участка прибрежья. Онегативном влиянии длительной задержки ледовых массивов на условия обитания гидробионтов можно косвенно судить потому факту, что в этой зоне прибрежья отсутствуют постоянные нерестилища гижигинско-камчатской сельди.
Антициклонические микроциркуляции характеризуются слабым течением и способствуют удержанию молоди в пределах заливов и равномерному распределению кормового планктона по их акватории. Распреснение водантициклонических вихрей благоприятно сказывается на смолтификации молоди в прибрежье и накоплении биомассы кормового мезопланктона, препятствует проникновению хищных форм планктона, способных вступать в пищевую конкуренцию с молодью горбуши. Еще одна положительная для выживания молоди черта антициклонических гидродинамических образований заключается в заглублении теплых поверхностных вод, что способствует раннему и быстрому прогреву водных масс.
Таким образом, циклонические водовороты, в отличие от антициклонических, не создают благоприятных условий для успешного выживания горбуши, что сказывается на величине ее запасов в реках того или иного участка побережья. Как следует из табл. 42, Ольская и Гижигинская группы рек, обладающие наибольшими запасами горбуши, по своим границам совпадают сантициклоническими вихрями, тогда как Тауйская и Ямская группы рек, обладающие существенно меньшими запасами, — с циклоническими циркуляциями.
Кроме того, разница в условиях раннего морского периода жизни в западной и восточной частях Тауйской губы позволила горбуше Ольской группы pек в середине 1980-х гг. перейти на доминирование по ряду поколений четных лет. Особенностью этого перехода является то, что четный ряд поколений горбушихарактеризуется меньшей плодовитостью и, соответственно, меньшим репродукционным потенциалом по сравнению споколениями ряда нечетных лет (табл. 43).
Таблица 43. Биологическая характеристика горбуши северного побережья Охотского моря*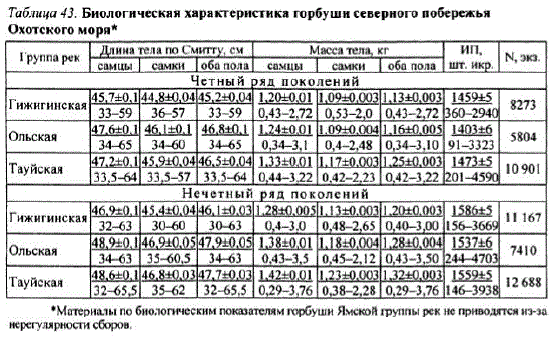
"Материалы по биологическим показателям горбуши Ямской группы pек не приводятся из-за нерегулярности сборов,
Соответственно, для того, чтобы генерация четного ряда лет могла стать доминирующей, в прибрежье должны сложиться исключительно благоприятные условия. Согласно данным Г.В. Хена (1991), основанным на изучении ледовитости Охотского моря, с 1984 г. в Охотском море наступил теплый период. Это стало отправной точкой для перехода горбуши Ольской группы pек надоминирование почетной линии лет. Наступление в конце 1990-х гг. холодного периода привело к возврату динамики численности горбуши этой группы pек на доминирование поколений ряда нечетных лет с 2000 г.
На основании изложенного можно заключить, что выделенные математическимметодом группы рек реально существуют. Помимо отличий в динамике численности, они с достаточным уровнем достоверности (p<0,05) различаются практически во всех случаях по основным биологическим показателям (табл. 44).
Обращает на себя внимание тот факт, что горбуша Гижигинской группы рек имеет наименьшие, а горбуша Ольской и Тауйской — наибольшие размерновесовые показатели. Кроме того, горбуша соседних групп рек — Ольской и Тауйской — имеет очень близкие, но, тем не менее, достоверно различимые размерно-весовые показатели.
Таблица 44. Сравнение горбуши выделенных групп рек с помощью критерия Стьюдента
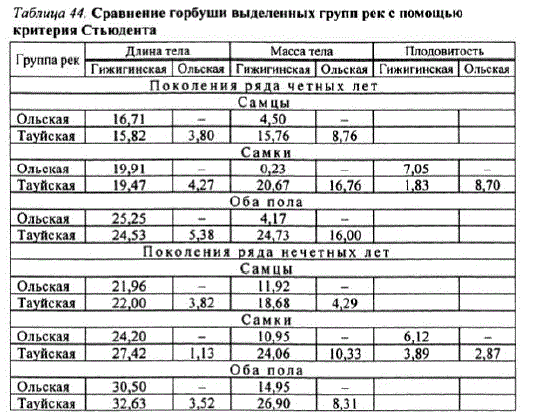
По нашему мнению, горбуше выделенных групп рек, согласно номенклатуре группировок внутривидового ранга, больше всего подходит статус локальных стад: они различаются динамикой численности и биологическими показателями, а в период размножения занимают определенную территорию (Иванков, 1993, 1997).
Темпоральная подразделенность. В настоящее время имеются многочисленные данные осуществовании сезонных расгорбушина значительной части eе ареала (Иванков, 1967а, б., 1971, 1991, 1993, Гриценко, 1981, Гриценко H np., 1987; Skud, 1958; Vernon, 1962; Heard, 1991; H III,p.).
Северное побережье Охотского моря до сих пор остается слабоизученным в этом отношении. Сведения о неоднородности горбуши в водоемах этого участка ареала носят предварительный характер (Ионов, 1987, Марченко, 1999). Главная сложность, возникающая при выявлении подобных структур, — значительное перекрывание сроков нерестового хода сезонных группировок.
Первоначально мы попытались выявить сезонные группировки горбушив одной из самых северных рекзал. Шелихова — Гижиге. За исключением разрозненных данных по срокам нерестового хода и некоторым биологическим показателям (Кагановский, 1949; Волобуев и др., 1998), какие-либо другие сведения о гижигинской горбушев литературе отсутствуют.
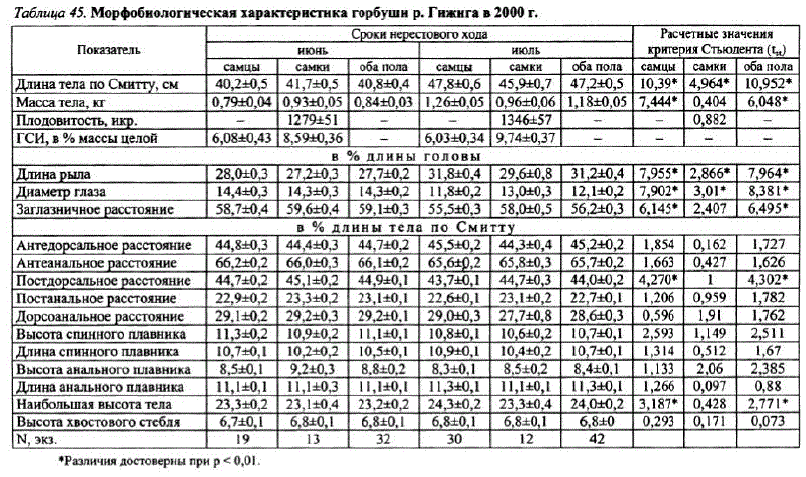
Материал собран во время экспедиционных работ на р. Гижига в 2000 г. Из-за малочисленности четного поколения горбушир. Гижига и сильной разреженности ее нерестового хода были проведены только 3 морфометрических анализа (74 экз.), характеризующих нерестовую горбушу в период с начала до середины хода. Дальнейшие сборы были прекращены из-за эпизодичности и нерегулярности поимок производителей.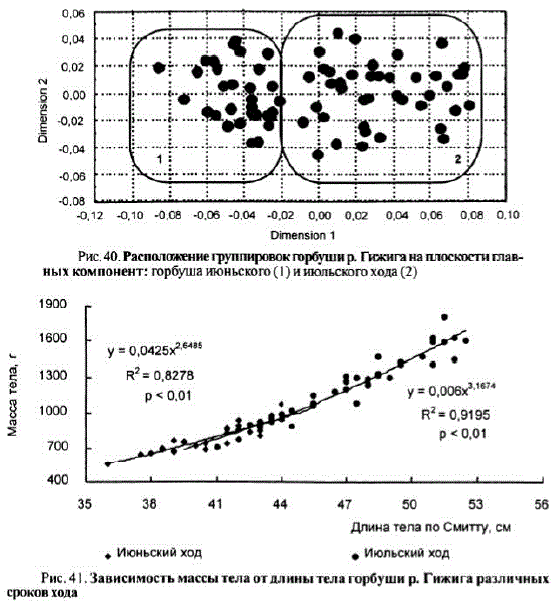
Рис. 41. Зависимость массы тела от длины тела горбушир. Гижига различных сроков хода
При изучении популяционной организации лососей большое внимание уделяется исследованию структуры чешуи и темпа роста (Бивен, 1961; Иванков, 1967а, б, 1993; Куликова, 1970, 1975; Грачев, 1983; Николаева, Семенец, 1983; Бугаев В.Ф., 1995; Иванков, Бисага, 1995; Каев, 1998; Кузищин и др., 1999).
На чешуйной пластинке гижигинской горбуши июньского хода в среднем образуется 32,1 склерита, июльского — 33,9 (t = 3,29, p <0.01). Впервый годна чешуе горбуши обеих группировокзакладывается около 23,5 склерита(t = 0,64, p > 0,01). Разница в количестве склеритов на чешуе появляется в зоне прироста, где y июньской горбуши закладывается 8,7 склерита, а у июльской — 10,3 (t = 3,57, р < 0,01). Исходя из того, что темп образования склеритовугорбуши постоянен на протяжении всей жизни (Иванков, 1967б), можно с определенной долей уверенности заключить, что горбуша июльского хода проводит в море больше времени.
4.
Горбуша июльского хода имеет более высокий темп роста (t = 3,54, p<0,01). При практически равном количестве склеритов в первый год жизни на чешуе рыб обеих группировок июльская горбуша достигает 35,7 см, тогда как июньская только 33,6 см, что, возможно, свидетельствует о разных районах нагула в океане или о более длительном сроке нагула. Отличия есть также и в приросте за последний год жизни: для июльской горбуши он составляет в среднем 11,4 см, для июньской — 7,1 см (t = 6,29, p < 0,01).
По мнению ряда авторов, изменение соотношения полов, размерно-весовых показателей, коэффициента зрелости на протяжении анадромной миграции свидетельствует о проходе на нерестилища разных экологических форм горбуши (Иванков, 1967а, б; Гриценко и др., 1987; Heard, 1991; и др.). Например, в реках Южных Курил воспроизводятся две формы горбуши — ранняя и поздняя. Как и в нашем случае, первая имеет небольшие размерно-весовые показатели, ее характерной особенностью является то, что самки крупнее самцов. Вторая характеризуется большими размерами и плодовитостью; у нее, как и у подавляющего большинства других популяций горбуши, самцы крупнее самок (Иванков, 1967б).
Проведя аналогии, можно сделать предположение о существовании угорбушир. Гижига двух экологических форм: ранней — совершающей нерестовую миграцию в июне, и поздней — идущей на нерест в июле. Как уже говорилось, эти экологические группировки статистически достоверно различаются сроками нерестового хода, биологическими показателями, темпом роста, структурой чешуи. Анализ многолетнего материала, накопленного лабораторией по изучению лососевых рыб МагаданНИРО, показал, что вр. Гижига случаи, когда в начале нерестового хода в июне самки были крупнее самцов, наблюдались и ранее (1972 г.). Такая же картина имела место в реках Тауйской губы — Оле (1975, 1984, 1985, 1998, 1999) и Тауе (1975, 1982). Эпизодичность появления в сборах горбуши раннего (июньского) хода, с одной стороны, объясняется совпадением сроков ее миграции с весенними паводками, существенно ограничивающими возможность сбора материала, с другой — ее крайнемалой численностью.
Все это косвенно указывает на неоднородность горбушине толькор. Гижига, но и всего северного побережья Охотского моря и подтверждает опубликованные ранее данные (Марченко, 1999).
Смотрите также






