Экология и биология молоди кеты в пресных водах
Покатная миграция молоди тихоокеанских лососей из мест рождения в реках Северо-Востока Азии в прибрежные морские акватории является важным этапом их жизненного цикла. Контроль за уровнем покатной миграции молодилососей в реках материкового побережья Охотского моря позволяет оценить эффективность естественного воспроизводства лососей. Ежегодные мониторинговые работы по количественной оценке и качественному составу молодикеты в период ее ската в море позволяют получать сведения о ее относительной численности и биологических показателях. Сопоставление полученных данных с имеющимися за предыдущие годы дает возможность оценить уровень выживаемости и биолого-физиологический статус покатной молоди. Данные об уровне ската молодикеты, получаемые на модельных водоемах, используются в качестве оценки начальной численности ее генераций. Показателем выживаемости являются коэффициенты возврата от скатившейся молоди для малочисленных, средней численности и многочисленных поколений, которые используются в прогнозных оценках.
Период адаптации в эстуарно-прибрежной зоне и переход в воду морской солености сопровождается гибелью значительной части выжившей в пресных водах молодилососей. Считается, что этот этап в жизненном цикле тихоокеанских лососей является одним из наиболее уязвимых. По оценкам ряда исследователей, смертность молоди отразличных стрессов (низких температур, нарушения механизмов осморегуляторной адаптации, высокого уровня ледовитости в прибрежье, слаборазвитой кормовой базы, аномальных паводков и др.), хищников и болезней в морской и океанический периоды жизни может достигать 95-98% (Neave, 1953; Foerster, 1955) HUI H azke 98-99% (UleBaH HILoB, 1969). B связи с этим в последние годы в эстуарии и прибрежье основной лососевой реки Тауйской губы (р. Ола) проводятся работы по оценке выживаемости, распределения, осморегуляторной адаптации и физиологического статуса молоди кеты перед ее выходом в прибрежье и откочевкой в открытые районы моря (K43eprm Ha, M3eprHH, 2008, 2009; Izergina et al., 2006).
Тем не менее информацию о сроках, суточной ритмике и динамике покатной миграции, показателях биологической структуры молоди считаем важной и необходимой для использования при проведении других работ, связанных с оценкой распределения, миграций и выживаемости молодикеты.
Начало ската молодикеты совпадает с наступлением весеннего паводка в реках материкового побережья Охотского моря (обычно это середина — конец II декады мая). Ежегодно проводимые на рр. Наяхан, Яма, Ола, Тауй, Челомджа, Кухтуй, Охота работы по оценке численности покатной молодикеты показали, что стимулирующими факторами, определяющими ее миграционное поведение, являются увеличение скорости течения, подъем уровня и снижение прозрачности воды. Пик покатной миграции позднейкеты обычно приходится на начало — первую половину июня. Скат основной массы молоди заканчивается в конце июня. Слабая миграционная активность наблюдается в июле. Единичные экземпляры молодикеты попадаются в уловах малькового невода в августе и даже в сентябре в нижнем течении нерестовых рек.
В течение суток в реках побережья наибольшее количество молодикеты скатывается в темное время — с 0 до 3 ч, менее интенсивно скат продолжается до 6-7 ч утра. В ночные и утренние часы скатывается до 80—90% суточной численности покатной молоди. Разреженный скат наблюдается в дневное время, особенно в пасмурные дни и сподьемом уровня воды. Однако известны случаи, когда молодь мигрировала из реки днем (Костарев, 1970а). По данным
этого автора, основной скат молодикеты в нерестовой р. Улхан (Охотский район), являющейся притоком крупной нерестовой р. Кухтуй, приурочен к ясными безоблачным дням с пиком миграции в утреннее время, тогда как в самой р. Кухтуй основная масса молодискатывается в ночное время.
Дневной скат молодикеты отмечен из рек Приморья (Горяинов и др., 2007). Большая часть молоди скатывается с 12 до 16 ч. Напротив, меньшая часть молоди мигрирует в ночное время — с 23 до 2 ч. Скат основной массы молоди происходит в I—II декаде апреля.
Время ската молоди кеты на азиатском и североамериканском побережье Тихого океана зависит от климатических условий, протяженности рек, наличия внутривидовых форм. В северных районах Дальнего Востока (р. Анадырь) покатная миграция кеты начинается в конце мая—июне (Черешнев и др., 2002), в южных районах (южные Курилы) — с конца апреля — начала мая (Каев, Струков, 1999; Каев, 2003), на о. Хоккайдо — в марте (Sano, 1966; Kobayashi, Kurohagi, 1968). В малых реках скат кеты проходит за один месяц, а в таких крупных водоемах, как Юкон, он может продолжаться от вскрытия рек до сенTA6ps (Martin et al., 1986).
О.Ф. Гриценко с соавторами (1987) для рек Сахалина также отмечает преимущественно ночной скат молодикеты с пиком в 24 ч. Скат происходит с начала мая до начала августа. Длительность ската объясняется миграцией молоди двух раскеты — летней и осенней. Авторы отмечают связь динамики покатной миграции кеты с температурой воды: согласно их данным, с повышением температуры интенсивность ската молоди увеличивается. Максимум ската отмечен при температуре воды 7-8°С. Другие авторы (Воловик, 1967а,б; Костарев, 1970а, Шершнев, Жульков, 1979) считают что колебания температуры воды не влияют на интенсивность ската молодилососей. На Курилах интенсивность ската молодикеты определяется только уровнем воды (Чупахин, 1975).
Длительность покатной миграции зависит от внутривидового экологического разнообразия кеты, воспроизводящейся в бассейне реки. На Сахалине молодь кеты разнокачественна по составу, выделяются две экологические группировки: первая — с преобладающими размерами 27—40, вторая — 40—54 мм, поэтому скат ее длится до 3 мес (Гриценко и др., 1987). Две группировки покатной молодикеты отмечены и для Амура (Рослый, 2002): одна представлена относительно мелкой молодью с массой тела 240—320 мг, вторая — крупной молодью, с массой тела до 1,5—2,0 г. В бас. р. Тауй на побережье Охотского моря вследствие того, что в ней размножается кета двух экологических группировок, различающихся по времени захода на нерест, покатная миграция молоди продолжается с середины мая по конец июля и также выделяются две группировки молоди.
По всему ареалу размножения кеты суточный ритм ската ее молодисходен и зависит в основном от освещенности и прозрачности воды. Как правило, миграция молоди с нерестилищ вниз по течению на всем ее ареале начинается с наступлением сумерек и заканчивается в утренние часы с рассветом. Пик ската приходится на самое темное время суток (Волобуев, 1984, Гриценко и др., 1987; Сметанин, 1991; Каев, Струков, 1999, Волобуев М.В., Волобуев, 2009: Neave, 1955; Hunter, 1959; McDonald, 1960; Kobayashi, Ishikawa, 1964, Koski, 1975; Martin etal., 1986; Salo, 1991; H up.).
Рассмотрим конкретные примеры динамики и особенностей ската молоди из основных нерестовых лососевых водоемов материкового побережья Охотского моря. Скат молодикеты в р. Наяхан на северо-востоке побережья происходит с конца мая — начала июня по конец июня — начало июля (рис. 18, a).
Пик покатной миграции обычно наблюдается в I декаде июня. Температура воды за период ската молоди плавно повышалась от 1 до 8°С. Связи интенсивности ската с температурой воды не установлено. В большей степени обнаруживается связь ската с уровнем воды.
В р. Ямаскат молоди происходит с конца мая — начала июня по конец июня — начало июля (рис. 18, б). Пик покатной миграции обычно наблюдается во II декаде июня. Всплески численности покатной молоди в виде малых пиков сопряжены, как правило, с подъемами уровня воды в реке.
Большая часть молоди кеты (до 60%) cкатывается в период с IV по VI пятидневку июня. В течение покатной миграции наблюдается постепенное повышение температуры воды от 0,2 до 13,0°C. Можно заметить, что интенсивность покатной миграции зависит от уровня воды в реке и слабо связана с темnepaTypon BoIHoro IIoToKa.
Скат покатной молоди кеты в р. Ола происходит с IV—V пятидневок мая по конец июня (рис. 18, в). Пик миграции приходится на 1 или II декаду июня, что зависит от погодных условий и паводкового режима реки. В динамике покатной миграции молодикеты р. Ола (2004 г.) наблюдались два пика численности: первый отмечен в I—II пятидневках июня, второй — в III—V пятидневках июня. За это время скатилось, соответственно, 37,6 и 60,8% молоди. Незначительная прямая взаимосвязь отмечена между динамикой покатной миграции молодии изменениями уровня и температуры воды. За период проведения работ температура воды в р. Ола возросла с 1,2 до 7,5°С.
Скат молоди поздней кеты в р. Tayй происходит с начала III декадымая T10 KOHell HIOHH (pHC. 18, 2).
Основной скатмолоди кеты в 2004 г. пришелся на III декаду мая — Ідекадуиюня, за это время скатилось 80% покатников. Между динамикой скатакеты и изменениями уровня и температуры воды была отмечена прямая слабая взаимосвязь: R = 0,3 и 0,4, соответственно, (p < 0,05). Температура воды за время проведения работ повысилась с 0,6 до 12,0°C.
В бас, р. Кухтуй (р. Улхан) (рис. 18, д) скат молоди происходит сначала июня по началоиюля. Пик покатной миграции обычно приходится на II—IIIдекады июня. Очевидно, скат молоди поздней формы кеты с нерестилищ в реках материкового побережья Охотского моря, как в северо-восточных, так и в югозападных районах, происходит с конца мая по конец июня — начало июля. Более ранние или более поздние сроки начала и окончания ската молоди сопряжены с погодными условиями и могут отклоняться в ту или другую сторону на неделю — 10 дней. Отмечена связь интенсивности ската молодикеты с подъемами уровня воды в реках. Зачастую ряд мелких пиков в динамике ската обусловлен колебаниями уровня воды. Связи уровня ската молоди с температурой невыявлено. Основная масса молодискатывается в темное время суток. Такие же данные были получены при проведении учета молодилососей на основном руслер. Кухтуй (Китов, 2005).
Основные показатели покатной молоди поздней кеты заряд лет приведены в табл. 16.
Длина тела по Смитту в среднем колеблется от 35 до 39 мм, масса тела — от 344 до 465 мг. Значительная часть покатной молоди (от 33 до 83%) питается — общий индекс наполнения желудков не превышает 200"looo. Следует отметить, что скат части молоди поздней формы (по экспертной оценке, до 15% покатников) происходит еще до ледохода и основного паводка. Это, кaк правило, крупная молодь, рано вышедшая из бугров и нагуливавшаяся на нерестилищах до подъема уровня воды в реке. То же самое отмечено и для р. Анадырь: около 12% молоди здесь мигрирует до основного ската подо льдом (Черешнев и др., 2001).
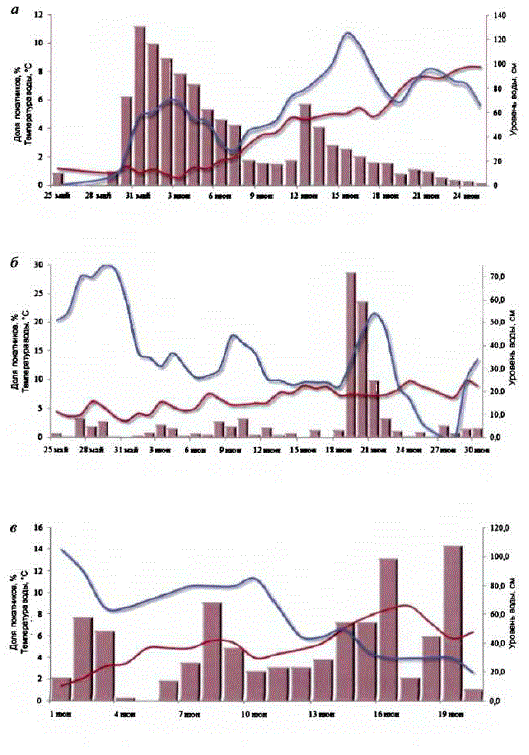
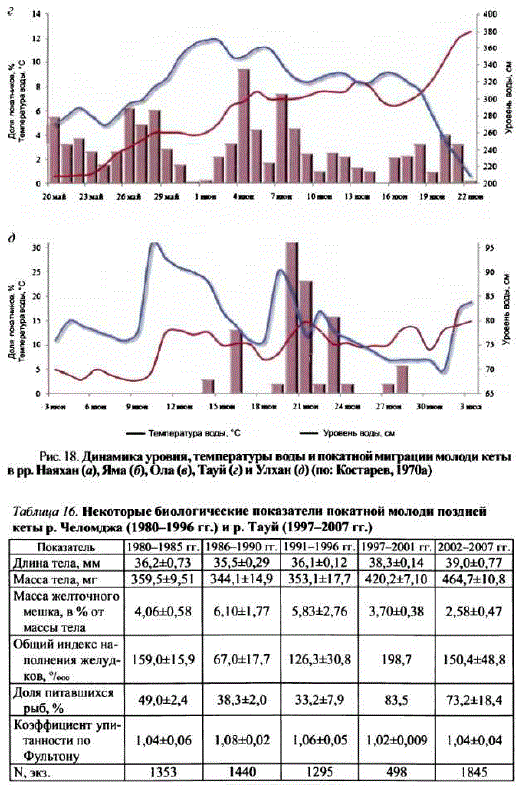
Рис. 18. Динамика уровня, температуры воды и покатной миграции молодикеты в рр. Нанхан(а), Яма (б), Ола (в), Тауй (г) и Улхан (д) (по: Костарев, 1970a)
Таблица 1б. Некоторые биологические показатели покатной молоди поздней
Сроки ската и биологические характеристики молоди ранней кеты могут отличаться в зависимости от типа и величины водоема. Так, например, скат молоди ранней кеты из рек равнинного типа, какой является р. Тауй с ее правым притоком р. Кава (протяженность 280 км), происходит с середины июня до середины июля.
Пик покатной миграции молоди ранней формы отмечается обычно в конце июня — I декаде июля. Основная масса молодискатывается в темное время суток. Средняя длина тела покатной молоди ранней кетыр. Тауй поколений 1996— 2001 гг. варьировала от 51,0 до 52,5 мм (48—63 мм), масса тела — от 1173 до 1378мг (836—2433 Mr) (Ta6JI. 17).
Таблица 17. Размерно-весовая характеристика покатной молоди ранней и поздней форм кеты некоторых рек материкового побережья Охотского моря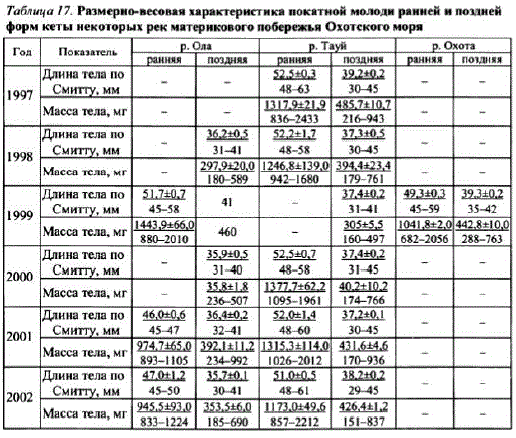
Вр, Ола молодь ранней кеты несколько мельче — ее средняя длина колеблется от 46 до 52 мм (45—58 мм), масса тела — от 945 до 1444 мг (833—2010 мг). По р. Охота данные о молоди ранней кеты имеются лишь за один год. Ее средняя длина и масса сходны с аналогичными показателями молоди из рр. Тауй и Ола.
На рисунках, приведенных в статье В.В. Китова (2005), также четковыделяются два пика численности мигрирующей молоди в р. Кухтуй: один до середины 20-х чисел июня — очевидно, это молодь поздней кеты, о чем свидетельствует небольшая масса тела рыб (300—400 мг). Второй — с 20-х чисел июня по начало — конец I декады июля (ранняя форма кеты с массой тела рыб 500—700 мг). Крупная молодь ранней кеты размером более 50 мм уже может иметь чешую. Число склеритов у таких рыб варьирует от 1 до 4. Размерновесовые показатели покатной молоди поздней кеты других рек сходны с таковыми для р. Тауй (см. табл.17).
84
Сроки, суточная динамика ската и биологические показатели молодиранней кеты могут не отличаться от таковых поздней формы. Так, в р. Мотыклейка(протяженность ее 49 км) молодь ранней кеты совершает покатную миграцию со II декады мая по конец июня, большая часть ее скатывается в темное время суток. Средний размер покатной молодимотыклейской кеты (N = 300 экз.) cocTaBHJI 35,6 MM (30—42 MM), Macca TeJIa — 266 MT (165—580 Mr).
Значительные размеры и масса тела молоди ранней кеты из крупных нерестовых водоемов обусловлены, по-видимому, особенностями воспроизводства кеты этой формы в бассейнах рек. Нерест ее происходит преимущественно в предгорных притоках 1—2-го порядка, значительно удаленных от устья. Так, например, в бас. р. Кава, скатываясь весной с нерестилищ, молодь попадает в основное русло, где медленное течение, хорошая кормовая база и высокая (до 15—18°C)температура воды. Река Кава играет роль выростного водоема для скатывающейся молоди ранней кеты. Продвигаясь вниз, она интенсивно питается веснянками, поденками, хирономидами и двукрылыми. В реках побережья молодь ранней кеты выходит из бугров раньше, чем молодь поздней формы, нагуливается на нерестилищах и питается во время покатной миграции, поэтому в период ската имеет гораздо большие размерно-весовые характеристики.
В малых реках, какой является Мотыклейка, молодь ранней кеты не имеет условий для длительного речного нагула и выносится весенним паводком сразу в морское прибрежье зал. Мотыклейский.
Очевидно, выживаемость молоди ранней кеты должна быть выше, так как она характеризуется более высокими размерно-весовыми характеристиками к моменту ската. В последние годы действительно отмечено увеличение доли ранней кеты в отдельных популяциях до 45—47% (Волобуев, Голованов, 2001), тогда как в 70—80-е гг. XX в. ее доля в подходах не превышала 5—6% (Евзеров, 1983). Рост численности ранней кеты, обусловлен, по-видимому, прекращением масштабного океанического дрифтерноголова и общим улучшением климатоокеанологической обстановки в экосистемах дальневосточных морей и в северо-западной части Тихого океана, что позволило восстановить внутривидовую структуру популяций.
В целом следует отметить, что скат мальковкеты поздней формы происходит в более сжатые сроки, тогда как скат молоди ранней формы может быть растянутым по времени и труднее поддается учету. Поэтому в качестве предикторов прогнозных оценок численности формирующихся поколений кеты, как правило, используются более полные данные о количественных показателях покатной миграции и коэффициентах возврата поздней кеты, которая в настоящее время является доминирующей экологической формой на материковом побережье Охотского моря. Однако учитываются имеющиеся данные очисленности и биологической структуре и ранней формы кеты.
Выживаемость кеты от откладывания икры в грунт доската молоди в море определяется влиянием комплекса абиотических и биотических факторов. Абиотические факторы основное влияние оказывают в период нереста, эмбриогенеза и в эстуарно-морской период жизни; воздействие биотических факторов происходит в основном после выходаличинок из бугров и перехода их на экзогенное питание (Костарев, 1964б, 1970; Леванидов, 1969, Рослый, 1970; Гриценко и др., 1987; и др.). Влияние абиотических факторов сопряжено с особенностями климатических условий. Их роль и изменения обусловлены сезонными и долгопериодными колебаниями климата. Воздействие абиотических факторов на воспроизводстволососей начинается с захода их в реки. В период анадромной миграции, нереста, инкубации икры иската молодилососей основными факторами, определяющими эффективность воспроизводства лососей,
85
являются гидрологический и метеорологический режимы в районе нерестовых водоемов. Выживаемость кеты сильно колеблется погодам в зависимости от конкретных условий воспроизводства: водности рек в летне-осенний период, сроков и высоты образования снежного покрова, температуры воздуха, обсыхания и промерзания нерестилищ, выпадения осадков.
Большое значение имеет водность рек, которая обеспечивает проход производителей на нерестилища и обусловливает их распределение по бассейну. В годы смощными паводками уменьшается промысловая нагрузка на популяции и на нерест проходит большее количество производителей, и, наоборот, в маловодные годы повышается интенсивность промыслового изъятия и ограничивается доступность нерестилищ. Повышенные уровни и паводковые воды снижают эффективность нереста, так как частично нерест при повышенных уровнях воды происходит за пределами оптимальных границнерестилищ и после спада воды они обсыхают, а с наступлением холодов промерзают. Кроме того, в паводки типичные многолетние нерестилища становятся малопригодными для нереста, поскольку скорости течения на них возрастают и много икры — до 50% теряется вследствие вымывания ее из гнезд (Леванидов, 1965). Гибель амурской кеты в среднем составляет около 50% от средней абсолютной плодовитоcти (Кузнецов И.И., 1937). По данным В.Н. Иванкова (1968), на Курилах потери икры при нересте составляют от 20 до 36% (33,6). За период инкубации гибнет в среднем около 43,5% икры от заложенной в грунт. По данным А.М. Каева (1980), выживаемость южнокурильской кеты превышает 80%. По мнению автора, такая высокая выживаемость объясняется высоким дебитом грунтовых вод, на выходах которых размножается кета.
Особенно сильно колебания уровня воды в реках воздействуют на выживание икры в зимний период за счет обсыхания, промерзания нерестовых площадок, а также в результате снижения проточности и заиливания нерестовых бугров. Кроме того, паводки, проходящие после массового нереста, разрушают нерестовые бугры. Динамика водозапаса и уровень дебита грунтовых италиковых вод во многом зависят от времени выпадения снега и высоты снежного покрова, так как своевременно выпавший снегиграет роль теплоизолятора и способствует более равномерному расходу грунтовых вод в течение зимы. В малоснежные, с низкими температурами зимы происходит глубокое промерзание почвы, в результате чего отмечается промерзание таликовых образований, насыщенных водой, и снижается или вовсе прекращается ее поступление из водоносных горизонтов. Между высотой снежного покрова в декабре и величиной потомства, полученного от одной самки, установлена высокая степень корреляции — 0,79 (Костарев, 1970г). Таким образом, температурный фактор в сочетании с высотой снежного покрова существенно влияет на выживаемость икры иличинок кеты. В зависимости от степени влияния перечисленных факторов колеблется и выживаемость потомства лососей. Среднемноголетний коэффициент скатакеты в реках юго-западной части побережья колебался от 0,3 до 35,5%, составив в среднем 15,7% (Костарев, 1970б). В реках северо-восточного участка показатели выживаемости гораздо ниже — коэффициент ската колеблется в пределах 2,1—9,6%, что, возможно, обусловлено более суровыми климатическими условиями региона. Среднемноголетний коэффициент возврата североохотоморской кеты от отложенной икры составляет 0,11—0,13%, кеты западного участка побережья — 0,13% (Костарев, 1970б). Коэффициент возврата от скатившейся молодикеты рек охотского стада составил 1,9% при колебаниях от 0,69 до 5,16% (Костарев, 1970б). В реках северо-восточного участка побережья (тауйское, ямское, гижигинское стада) среднемноголетний коэффициент возвратабли
зок к 1,0% при колебаниях от 0,3 до 2,4%. Коэффициент возвратаамурской летней кеты в среднем составляет 1,6—1,7, осенней — 1,9% (Рослый, 2002).
В реках материкового побережья Охотского моря основными видами рыб, уничтожающими молодь тихоокеанских лососей, являются хариус, голец и кунджа. Самый массовый вид — сибирский хариус. Наблюдения, проведенные на pр. Охота, Челомджа, Яма и Гижига в 70-80-е гг. XX в., показали, что икра и молодь лососей служат важными компонентами питания факультативных хищников в мае-июне во время ската молоди с нерестилищ и в руслах рек на пути ее к морю. Частота встречаемости молодикеты и горбуши в желудках хариуса составила 48—100, гольца — 17—96, кунджи — около 1% (Волобуев, Кузищин, 1988). В предпокатной и покатной периоды молодь кеты в реках охотоморского побережья интенсивно поедается сибирским хариусом, который выступает в роли основного факультативного хищника пресных вод. В конце мая—июне хариус в районе нерестилищ питается в основном покатной молодью кеты, которая составляет 78—95% от массы пищевого комка. В основном он образует массовые скопления в устьях нерестовых проток, где интенсивно поедает скатывающуюся с нерестилищ молодь. Максимальное значение индексов наполнения желудков при питании молодью лососейдостигалось к пику ее покатной миграции, которая обычно приходится на конец мая — начало июня. В желудках хариуса встречено до 120—170 экз. молодикеты. В среднем за годы наблюдений (1978—1992) количество мальковкеты на один желудок колебалось от 10 до 42 экз. Частные индексынаполнения желудков были высокими—256—343°loco (Волобуев, Рогатных, 1999). Следует отметить, что вторым позначимости факультативным хищником в реках материкового побережья является голец проходной (SalveIітиs таІта). В желудках крупных гольцов (45—50 см) насчитывалось до 330, в среднем 12,6 экз. молодикеты. Сходный уровень потребления молодилососей гольцом для рр. Кухтуй и Охота приводят К.И. Попов (1958) и В.Л. Костарев (1970а). Индексы наполнения желудков угольца молодью кеты (до 170 экз. молодикеты и горбуши в одном желудке) колебались от 60 до 570°looo. Однако численность гольца, остающегося после зимовки в реках, к моменту ската молоди невелика, поэтому значительного негативного влияния на выживаемость и численность молодилососей он не оказывает.
Значительное выедание мальмой молоди горбуши показано для прибрежья северо-востока Камчатки (Тиллер, 2007). В желудках гольцов количество съеденной молоди горбуши достигало 400—940 экз. при индексе наполнения желудков до 1000°/boo.
Кунджа (S. Iеисотаетіs) cкатывается в море после зимовки в течение апреля — начала мая, поэтому в период ската молодилососей в реке она практически отсутствует и как факультативный хищник негативного влияния на формирующиеся поколения лососей не оказывает. В период нагула в морском прибрежье молодь лососей в ее питании встречалась редко и в небольшом количестве (в среднем менее 1 экз./желудок, средний частный индекс наполнения ее желудков молодью составил 2,18°/ooo (Волобуев, Никулин, 1975)). Основное значение в питании кунджи имели морские рыбы — навага и корюшка с наполнением желудков до 323°/ооо,
В реках Сахалина кунджа интенсивно поедает молодь лососей (Воловик, Гриценко, 1970), в озерах Южного Сахалина она в основном потребляет песчанку, корюшку и гольяна (Савваитова, 1964). В Приморье кунджа в значительном количестве поедает покатную молодь горбуши (Василенко-Лукина, 1962). Отмечено, что в годы с маловодными, непродолжительными паводками доля молоди, поедаемой факультативными хищниками, увеличивается, и, наоборот, чем сильнее и продол
жительнее весенний паводок,тем большеезначение в их питании приобретаетсносимый бентос. Отметим, что в маловодные весенние паводки сумеренной мутностью воды молодь доступна хищниками в основном русле, тогда как в годы с сильными паводками и повышенной мутностью воды хариус и голец вынуждены уходить с речного русла и отстаиваться в затонах и заливах на участках со светлой водой. При этом основная масса молоди выходит из-под пресса факультативных хищников и беспрепятственно преодолевает речной отрезок пути в период миграции в море (Фроленко, 1983). По мнению некоторых авторов (Воловик, Гриценко, 1970), факультативные хищники поедают в основном недоразвитую, нежизнеспособную молодь. Однако регулярно проводимый нами внешний осмотр хорошо сохранившейся молоди, вынутой из глотки и верхнего отдела желудка хариусов и гольцов, показал, что она была нормально развитой, хорошей упитанности, без желточных мешковикаких-либо признаков уродств, т.е. вполнежизнеспособной.
Кроме того, следует отметить, что большое значение на выживаемость в пресных водах производителей, эмбрионов и мальковкеты имеют и другие факторы биотического порядка: рыбоядные птицы, хищные млекопитающие и грибок Saprolegnia sp. (Волобуев, Кузищин, 1988). Некоторое негативное влияние на общую численность покатной молоди оказывают такие рыбоядные птицы, как крачки и речные чайки, многочисленные сезонные скопления которых ежегодно наблюдаются в весеннее время на лососевых нерестовых реках североохотоморского побережья. В течение светового дня они большую часть времени летают на низкой высоте над рекой, то и дело выхватывая скатывающуюся молодь из воды. По частоте охотничьих нырков чаек косвенно можно судить об интенсивности ската молодилососей.
Значительное выедание молодикеты факультативными хищными рыбами пресноводного комплекса (ленок, налим, хариус, гольян, голец) отмечено И.И. Кузнецовым (1937). Наибольший ущерб вследствие хищничества наносят ленок (до 35 экз. молоди в желудках), налим (до 43 экз. молоди) и гольцы (более 300 экз. молодикеты). По мнению этого же автора, огромный ущерб численности формирующимся генерациям лососей причиняют рыбоядные птицы, например, крачки. На Камчатке в районе Азабачьего озера за 2-месячный период во время ската, по подсчетам автора, они поедают не менее 1 млн крупных (5—6 см) мальковкеты.
Выедание хищниками пресных водскатывающейся молодилососей в бассейне Амура может достигать 22,5% от ее общего количества (Леванидов, 1969). Значительный ущерб в лимане Амура и морском прибрежье Сахалинского залива и Татарском проливе наносит выедание молодилососей миногой — до 50— 60% (Рослњи, 2002; Beamish, Roslу, 1995). Bболњшом количествемолодњлососей в лимане Амура и эстуариях камчатских рек (более 50%) поедается азиатской корюшкой (Леванидов, 1969; Карпенко, 1983).
Отрицательное воздействие на численность лососей во время их анадромной миграции оказывают настоящие тюлени — в основном это ларга, являющаяся доминирующим видом в прибрежье. В летнее время (июль-август) ларга повсеместно встречается вдоль всей береговой полосы (3—5 экз. на 1 км), но значительные скопления ее отмечены в приустьевых участках нерестовых рек. Численность таких скоплений зависит от величины реки и количества заходящих на нерестлососей. По экспертной оценке, численность ларги в пределах Тауйской губы составляет не менее 10 тыс. экз. (Задальский, 2004). В прилив ларгинаходятся в воде и охотятся за лососями, активно поедая их; во время отлива на косах и банках они образуют залежки иногда численностью донескольких тысяч. Рассчитанное теоретически возможное выедание лососей ларгой за сезон в прибрежье Тауйской губы составляет около 8 тыс. т.
Значительное количество нерестующих лососей поедается бурыми медведями, которые ко времени нереста концентрируются в районе нерестилищ. Особенно велико их влияние на лососей, преодолевающих перекаты и размножающихся на мелководных нерестилищах. В период нереста горбуши, кеты и кижуча на крупных нерестилищах охотоморских рек скапливается до 30—40 бурых медведей, которые в августе-сентябре питаются исключительно рыбой. По нашим наблюдениям, суточный рацион медведя в это время составляет 4— 6 лососей. Подсчеты показывают, что за нерестовый период только на одном контрольном нерестилище медведями уничтожается до 10—12 тыс. рыб. Помимо этого, медведи создают фактор беспокойства и нарушают нормальное течение процесса распределения по нерестовым участкам и нереста производителей (Волобуев, Кузищин, 1988). Значительное выедание бурыми медведямилососей на нерестилищах (до 75—100%) отмечено С.М.Коноваловым (1980), B.A. IlapeHcKAM (2005), E.B. Tony6b (2007).
В течение августа-сентября во время икрометания на нерестилищах лососей их икрой питаются хариус и голец. Накормленность хариуса икрой достигает 567°/ооо, гольца — 233°/ooo Частота встречаемости икры в желудках рыб достигала 100%. Однако ущерба воспроизводительному потенциалу популяции этим не наносится, поскольку поедается, как правило, потерянная при нересте икра, вымытая течением из бугров. Поэтому хариус и голец выступают здесь в роли биологических мелиораторов лососевых нерестилищ.
Значительное количество оплодотворенной икры кеты и кижуча гибнет от грибка Saprolegтia sp. в период инкубации в грунте нерестилищ - до 47-91% (Волобуев, 1984; Рогатных, 1987). Повышенная гибель от Saprolegтia sр. наблюдается в нерестовых буграх во второй половине зимы, когда снижается инфильтрационная способность грунтового потока в результате уменьшения водозапаса, аккумулированного в таликовых структурах. В это время в водоснабжении бугров увеличивается роль подруслового потока и с ним в гнезда попадают споры сапролегниевого грибка, который обильно поселяется на трупах производителей лососей, находящихся на нерестилищах.
Смотрите также






