Малоротая корюшка
Малоротая корюшка была описана Палласом в 1811 г. из водоемов Камчатки как Salmo olidus. В самостоятельный род Hypomesus малоротые корюшки выделены Жилем [Gill, 1863; приводится по Бергу, 1948]. Более чем полуторавековая история изучения этой группы рыб освещена в работе Мак-Аллистера [McAllister, 1963]. Значительный вклад в исследование систематики рода Hypomesus внесен в 60-е годы, когда, кроме упомянутой работы Мак-Аллистера, появились ревизии Хамады [Hamada, 1961] и В.А. Клюканова [1970],
каждая из которых существенно меняла сложившиеся представления о систематике малоротых корюшек. Эти исследователи, решая вопросы систематики, наряду с анализом данных по морфологии использовали сведения по экологии представителей рода Hypomesus. К началу наших исследований было описано пять видов малоротых корюшек [Клюканов, 1970], основные отличия которых в качестве своеобразного итога предыдущих исследований удобно представить в виде определительной таблицы:
1 (4) Позвонков 60 и более
2 (3) Нет рукоятки сошника, есть глубокий медиальный вырез в задней части mesethmoideum Н. pretiosus (Girard); морской прибрежный вид
3 (2) Есть рукоятка сошника, нет глубокого медиального выреза в задней части mesethmoideum Н. japonicus (Brevoort); морской прибрежный вид
4 (1) Позвонков менее 60 (50-59)
5 (6) Ductus pneumaticus соединяется с нижней стороной
плавательного пузыря, несколько отступая от его переднего конца Н. olidus (Pallas); пресноводный вид
6 (5) Ductus pneumaticus отходит от переднего конца плавательного пузыря
7 (8) Есть hypethmoideum и exethmoideum, нет рукоятки сошника Н. transpacificus (McAllister); предположительно проходной вид
8 (7) Нет hypethmoideum и exethmoideum, есть рукоятка сошника Н. nipponensis (McAllister); вид представлен проходной и жилой пресноводной формами
Наблюдаемое в природе многообразие форм малоротых корюшек и часто меняющиеся представления об их систематике побудили нас предпринять изучение рода Hypomesus. Вопросы систематики малоротых корюшек явились составной частью исследования их биологии, которое касалось также миграций, размножения, питания, роста и взаимоотношений с другими видами рыб, поэтому наши суждения о систематике малоротых корюшек в значительной мере основываются на экологических данных. Мы поставили целью выяснить таксономическое положение малоротых корюшек, обитающих на Дальнем Востоке, дать их подробное сравнительное морфометрическое описание, получить представление о географической и биотопической изменчивости.
Малоротые корюшки, обитающие на Дальнем Востоке, в наших материалах представлены тремя видами: Я. japonicus, H. nipponensis и Я. olidus. Ниже приводится их описание [Гриценко, Чуриков, 1983].
Н. japonicus. Наиболее крупная малоротая корюшка. Длина особей в нерестовых стадах 14-22 см. Распространена от Анадырского залива до Желтого моря (Чжифу), встречается на Сахалине, у южных Курильских островов и на Хоккайдо [Линдберг, Легеза, 1965]. Обитает в морской воде. Нерестится в апреле — мае на опресненных участках морских прибрежий с песчаным грунтом, при солености до 25 %о [Hamada, 1961], в ряде случаев заходит на нерест в лагуны с соленостью 4-6 %о. В I—II декадах мая отмечается ее массовый заход в озеро Тунайча, а в конце мая — в лагуну Буссе (Сахалин).
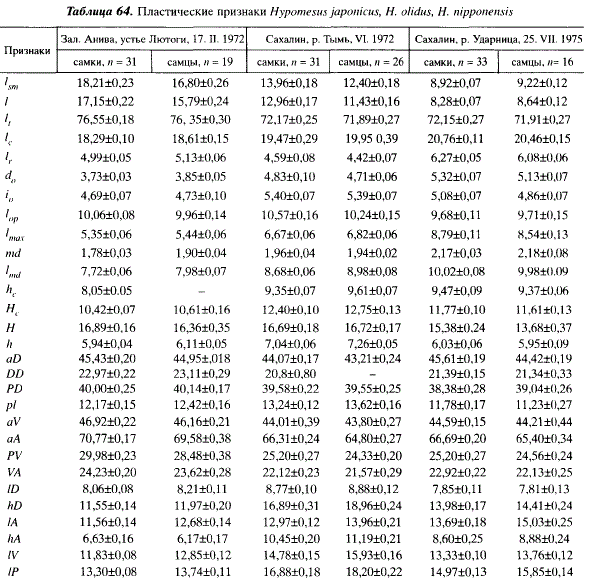
В пределах ареала Я. japonicus значения большинства меристических признаков (//; Р; A; D;p. арр) не обнаруживают закономерных изменений, связанных с широтным положением мест взятия проб или обитанием в бассейнах Японского и Охотского морей (табл. 63), однако в ряде случаев по некоторым признакам имеются достоверные различия между пробами, взятыми из близко расположенных мест. Например, корюшки из устья Лютоги и озера Тунайча различаются по числу пилорических придатков (td= 3,9; Р = 0,99). Различия подобного рода, видимо, связаны с существованием у Я. japonicus локальных популяций. В числе позвонков прослеживается слабо выраженная клинальная изменчивость. Значение этого признака у корюшки на севере ареала (Авачинский залив) выше, чем на юге (Амурский залив). Число жаберных лучей у корюшки восточного побережья Сахалина меньше, чем у корюшки западного побережья острова (td= 2,2; Р = 0,95). Число жаберных тычинок у корюшки из Японского моря меньше, чем у корюшки Охотского моря (td= 2,6; Р = 0,99). Сравнивая пластические признаки малоротых корюшек, мы видим, что Я. japonicus отличается от двух других видов более короткой и низкой головой, короткими челюстями, малым диаметром глаза и короткими плавниками, что характерно для морского пелагического вида. Расстояние PD; AV; AA; PV и VA у него короче, чем у других видов корюшек (табл. 64). У самцов 1А и IV больше, чем у самок, hA — меньше.
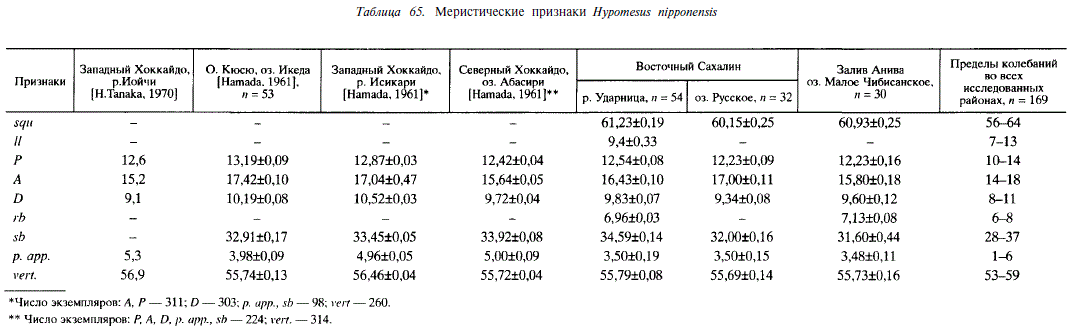
Н. nipponensis. Наиболее мелкая среди малоротых корюшек. Нерестовые стада ее состоят из особей длиной 8-13 см, однако рыбы длиннее 11 см встречаются исключительно редко. У этого вида нет специфичного огуречного запаха, столь характерного для двух других видов малоротых корюшек. Ареал Н. nipponensis простирается от Амура до Воньсана (Корея), эта корюшка встречается на южном Сахалине, острове Кунашир. В бассейне Амура встречена в Мергене, впадающем в озеро Чля, и в реке Джатта. В Японии известна на островах Хоккайдо и Хонсю, вселена в водоемы острова Кюсю [Клюканов, 1970; McAllister, 1963]. По сравнению с ареалами других видов малоротых корюшек ареал Я. nipponensis наиболее южный. Вид представлен проходными и жилыми (озерными и озерно-речными) формами.
По данным Хамады [Hamada, 1961] и нашим наблюдениям, Я. nipponensis — облигатный литофил, нерестится в ручьях и реках на перекатах и мелких плесах с галечно-песчаным грунтом. Озерные нерестилища расположены на мелководьях у берегов. Проходная форма обитает в Приморье, Японии и на южном Сахалине. В 1972 г. наблюдали особей, идущих из моря на нерест в реку Урюм (залив Анива). Малоротая корюшка, размножающаяся в реке Ударница, нагуливается в солоноватом озере Тунайча, хотя не исключено, что часть особей выходит в море. Эту форму корюшки в равной мере можно рассматривать как проходную и как озерно-речную. В Большом Чибисанском озере (южный Сахалин), соединенном с морем системой проток, обитает пресноводная форма Я. nipponensis. Примером типичной популяции из изолированных озер в наших сборах является корюшка озера Русское (южный Сахалин). Это пресноводное озеро — одно из группы озер, образовавшихся на месте морской лагуны, отделенное от моря мощной песчано-галечной косой. В озере имеются обширные галечные участки прибрежных мелководий глубиной 0,5-1,0 м, на которых нерестится Н. nipponensis. Интересно, что в соседних озерах такого же происхождения (Морж, Охотничье) из-за сильного зарастания нет подобной ситуации, и, как следствие, отсутствует Я. nipponensis. У особей данного вида отмечаются закономерные изменения ряда меристических признаков, связанные с широтным положением местообитания или типом водоемов (табл. 65). Так, у корюшки Японских островов больше пилорических придатков, чем у сахалинской (в среднем 4,85 ± 0,04 и 3,49 ± 0,09; Р 0,999; td= 13,9). При ранжировании выборочных средних значений этого признака в порядке возрастания обнаруживается, что оценки средних по выборкам Сахалина меньше, чем по выборкам из Японии, причем между наибольшим значением его на Сахалине (река Ударница) и наименьшим в Японии (озеро Икеда) различия достоверны (td= 2,3; Р = 0,95).
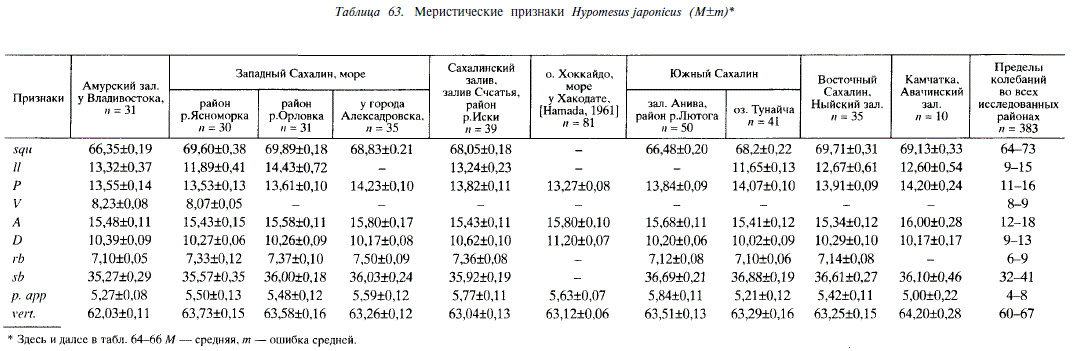
У малоротой корюшки из небольших озер (Икеда, Большое Чибисанское, Русское) число жаберных тычинок меньше, чем у проходной (река Исикари) или из крупных озер (Тунайча, Абасири) (в среднем 32,21±0,15 и 33,89±0,06; td= 9,8; Р = 0,999). Между наибольшим значением этого признака в первой группе рыб (Икеда) и наименьшим во второй (Исикари) имеется достоверное различие (td= 3,0; Р = 0,999). Обращает на себя внимание отсутствие различий в числе позвонков и пилорических придатков у корюшки из водоемов Тонино-Анивского полуострова (Сахалин).
По пластическим признакам Я. nipponensis отличается от двух других видов наибольшей длиной основания А, наибольшей длиной головы и челюстей, наиболее крупными глазами. Хвостовой стебель, PD, основание D наиболее короткие, высота хвостового стебля наименьшая. У самцов 1А и IP больше, чем у самок (Р = 0,99) (см. табл. 64).
Н. olidus. Согласно В.А. Клюканову [1970], этот вид распространен по азиатскому побережью, от реки Алазея (Восточно-Сибирское море) до Татарского пролива. На материковом побережье Японского моря Я. olidus отсутствует, а ранее сделанные указания ошибочны, так как за Я. olidus принимался, nipponensis. Е.И. Ивановой [1952] Я. olidus найден в районе Карской губы (озеро Круглое). По данным В.А. Клюканова [1970], Хамады [Hamada, 1961] и Мак-Аллистера [McAllister, 1963], это пресноводный вид. Однако Мак-Аллистер допускает возможность его выхода в солоноватую воду.
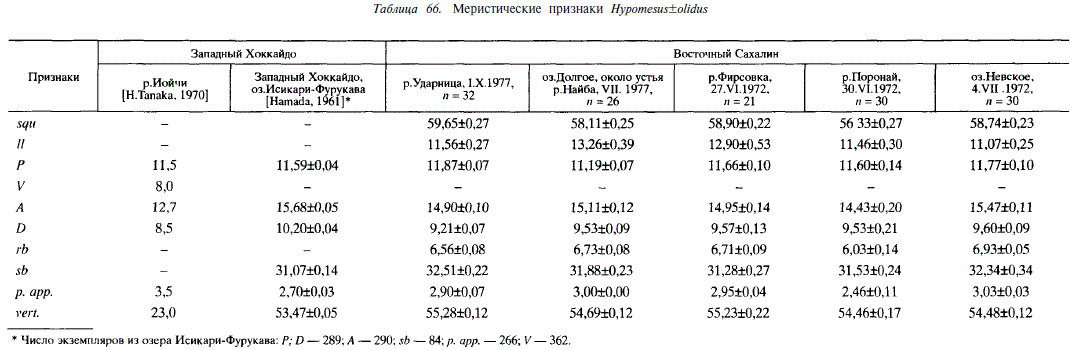
По нашим данным, Я. olidus представлен как жилыми (озерными и озерно-речными), так и проходной формами. Последняя весьма многочисленна в реках восточного Сахалина (Найба, Поронай, Набиль, Тымь, Даги, Фирсовка). В течение летних месяцев ее нагульные скопления в большом количестве отмечены нами в заливах Ныйский, Терпения; в солоноватоводном озере Невское, в море у поселка Стародубска, в Татарском проливе у города Александровска-Сахалинского и поселка Широкая Падь. Последнее обстоятельство дает основание включить северную часть Японского моря в ареал этого вида, тем более, что озеро Исикари-Фурукава и река Иойчи (остров Хоккайдо), где отмечен Н. olidus [Hamada, 1961; Н. Tanaka, 1970], также находятся в бассейне Японского моря.
Н. olidus из Ударницы, подобно Н. nipponensis из этого же водоема, может быть отнесен как к проходной, так и к озерно-речной форме. Типичная озерно-речная форма Н. olidus обитает в бассейне Амура, причем ряд популяций — на расстоянии 800-900 км от устья (озеро Болонь). К озерно-речной форме относится, судя по данным Белоусовой [1975], Н. olidus из озера Азабачье (Камчатка). Н. olidus обнаружена нами во многих изолированных пресноводных озерах Сахалина. В некоторых из них, относительно больших (Русское, Хвалисекое), этот вид обитает совместно с Н. nipponensis. В небольших озерах (Охотничье, Морж) обитает только И. olidus. Принадлежность малоротых корюшек из этих озер к пресноводной форме едва ли определяется наследственно, так как, по сути дела, это части проходных стад, отрезанные от моря физическими преградами. Время изоляции относительно коротко, так как озера Хвалисекое, Русское и Охотничье — водоемы молодые. О недавнем возникновении этих озер свидетельствует уже тот факт, что геологически молодым является их ложе — намытая морем песчаная коса. Связь с морем на протяжении истории подобных водоемов может возникать неоднократно как в периоды повышенной водности, так и при незначительных колебаниях уровня океана. На восточном берегу озера Русское мы обнаружили заросшее русло протоки, посредством которой через озера Донецкое и Айруп оно соединялось с морем, видимо, 100-200 лет назад.
Время изоляции в тундровых озерах, имеющихся в низовьях реки Тымь, из которых А.Я. Таранец [1937] описал подвид И. о. bergi, в принципе не может превышать нескольких столетий. Эти озера — давние старицы Тыми. Ложе их расположено в пойменном аллювии, на дне лежат остатки больших деревьев, обычно растущих по берегам сахалинских рек. Многолетний опыт работ в бассейне Тыми позволяет считать, что попадание паводковых вод в эти озера при экстремально высоком половодье вполне вероятно. Озера могут объединяться с рекой по меньшей мере раз в несколько десятилетий.
По нашим данным, Н. olidus в водоемах Сахалина, как и в водоемах Хоккайдо [Hamada, 1961], является фитофилом.
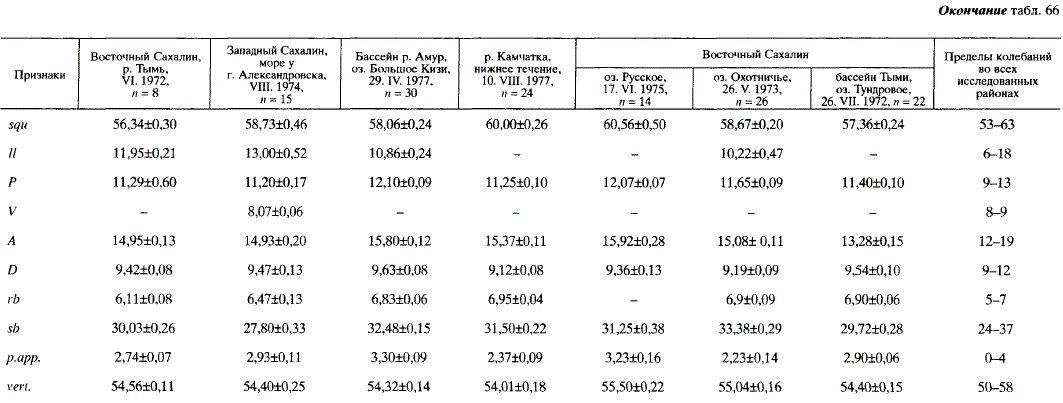
Анализ меристических признаков Н. olidus не обнаруживает закономерных изменений в связи с географическим положением населяемых водоемов или с принадлежностью к той или иной внутривидовой форме. Исключение составляет лишь число позвонков: у особей на южной границе ареала (Хоккайдо) их меньше, чем в других районах. По некоторым признакам в ряде случаев между корюшками из соседних водоемов существуют достоверные различия, вероятно, связанные с популяционной подразделенностью вида. Например, Н. olidus рек Фирсовка и Поронай достоверно различаются по squ; //; rb\ p. app (табл. 66).
Поскольку условия обитания проходной и живущей в пресных водоемах малоротых корюшек существенно различаются и, в первую очередь, это касается особенностей питания, было интересно провести сравнение по числу жаберных тычинок и пилорических придатков. С такой целью были вычислены средние величины этих признаков для двух суммарных групп. В первую вошли особи из озер Исикари-Фурукава, Кизи, Русское, Охотничье и Тундровое, во вторую — проходные особи, в том числе корюшки из солоноватоводных лагунных озер Невское и Долгое, служащих местом нагула проходной формы. Достоверных различий по этим признакам не обнаружено (среднее число жаберных тычинок равнялось соответственно 31,50 ± 0,12 и 31,28 ± 0,13; td= 1,27; пилорических придатков — 2,75 ± 0,03 и 2,78 ± 0,03; td= 0,75, что свидетельствует об их значительной стабильности. В связи с этим упразднение Мак-Аллистером [McAllister, 1963] подвида Н. о. bergi представляется вполне обоснованным.
По пластическим признакам проходная форма Н. olidus отличается от других видов наиболее широкой и высокой головой, массивным хвостовым стеблем, большими длиной и высотой спинного плавника и высотой анального, более длинными грудными и брюшными плавниками (см. табл. 64). Подобные особенности строения объясняются спецификой условий существования. Проходная корюшка совершает относительно длительную нерестовую миграцию, поднимаясь против течения на несколько десятков километров, что обусловливает необходимость мощного движителя. Откладывая икру на растительность, Н. olidus в большей мере, чем два других вида, совершает вертикальные перемещения в толще воды, в связи с чем необходимы более крупные плавники. Самцы и самки достоверно различаются длиной плавников: у самцов hD; IA; IP; IV и hA больше, чем у самок (Р = 0,95).
Таким образом, проведенные исследования подтверждают результаты ревизии рода Hypomesus, проведенной В.А. Клюкановым [1970], и дают основания считать, что принятая в настоящее время система этого рода соответствует ситуации, реально существующей в природе.






