Голец Salvelinus alpinus complex (Linnaeus)
Род Salvelinus (Richardson, 1836) подразделяется рядом исследователей на три подрода: Salvelinus (типовой вид S. alpinus, типовое местообитание Шведская Лапландия); Cristivomer (соответственно S. namaycush, бассейн Гудзонова залива); Baione (S.fontinalis, Лонг Айленд, Нью-Йорк) [Behnke, 1980 и др.].
Гольцы рода Salvelinus являются классическим образцом "трудного объекта" систематики. Большое разнообразие форм гольцов, населяющих водоемы разного типа, происхождения и геологического возраста, среди которых множество лабильных образований, находившихся в последние тысячелетия и десятки тысячелетий под воздействием оледенений и колебаний уровня моря, дает основание для разных, подчас противоречивых суждений.
Если в начальном периоде изучения гольцов эти противоречия в значительной мере порождались отсутствием информации, то в настоящее время их питают как отсутствие, так и наличие большого объема полевых наблюдений, экспериментов, а также применение новых методов исследований. В этой связи показательно замечание одного из наиболее компетентных специалистов, Р. Бенке [Behnke, 1980], который в конце своего обзора систематики рода Salvelinus написал, что "будет не шокирован, а скорее слегка удивлен, если в дальнейшем окажется, что большинство сделанных им таксономических построений не будет иметь тесного сходства с эволюционной реальностью".
Существует два подхода к таксономической оценке многообразия форм гольцов. При одном из них каждая форма в случае обнаружения морфологических различий с соседними формами или установления ее репродуктивной обособленности выделяется в качестве самостоятельного таксона [Викторовский, Глубоковский, 1977; Глубоковский, 1977а, б, в; Викторовский, 1978б; Черешнев, 1978, 1979; Викторовский и др., 1981; Глубоковский, Черешнев, 1981].
Представители второго направления, которое наиболее полно разработано К.А. Савваитовой, ее учениками и коллегами, считают, что многообразие форм гольцов Европы, Арктики и бассейна Тихого океана, относимых к видам S. alpinus (L.), S. maltna (Walbaum), S. lepechini (Gmelin), S. drjagini Logaschov, S. tolmachoffi Berg, S. jacuticus Borisov, S. czerskii Drjagin, S. toranetzi Kaganovsky, S. kronocius Victorovsky, S. albus Glubokovsky, S. neiva Taranetz, S. boganidae Berg, S. taimyricus Michin, S. andriaschevi Berg, следует рассматривать в рамках комплексного вида 5. alpinus complex [Барсуков, 1960; Савваитова, 1961а, б, 1979; Медведева, 1977а, б; Савваитова, Волобуев, 1978; Волобуев, 1978; Медведева-Васильева, 1978; Васильева, 1978, 1980; Михайлова, 1981; Savvaitova, 1980; Mednikov et al., 1980; Medvedeva, Savvaitova, 1980]. На основании анализа средних значений числа пилорических придатков и жаберных тычинок — признаков, широко используемых в систематике гольцов, К.А. Савваитовой и В.В. Волобуевым [1978] в пределах S. alpinus complex выделено три морфотипа: альпиноидный, мальмоидный и высокоарктический. Эти морфотипы распространены повсеместно, но преобладают соответственно в Европе, Арктике и бассейне Тихого океана. В пределах морфотипов существуют формы, различающиеся по характеру питания, степени оседлости, местам и срокам размножения, степени репродуктивной обособленности. Сахалинская мальма, в частности, рассматривается в качестве элемента мальмоидного морфотипа. Возникновение в разных участках ареала сходных по ряду признаков, но не тождественных фенотипов гольцов объясняется этими авторами как проявление гомологической параллельной изменчивости [Вавилов, 1967], а большинство симпатрических группировок рассматриваются в качестве многообразных форм адаптивного полиморфизма, которые не всегда следует принимать за начало симпатрического видообразования.
По определению М.В. Мины [1986], S. alpinus complex относится к категории полнокомплексных видов, т.е. включающих в себя как географически разобщенные, так и симпатрические группировки.
Ихтиологи Канады и США занимают позицию, среднюю между описанными выше, что, возможно, объясняется большей изученностью гольцов Северной Америки. Ими также выделяется в пределах рода Salvelinus небольшое число видов, но признается видовая самостоятельность S. malma, которая подразделяется на два подвида (северный и южный), различающихся числом позвонков [McPhail, 1961; Behnke, 1980; Cavender, 1980; Morrow, 1980; Johnson, 1980]. Р.Бенке рассматривает S. malma в качестве вида в пределах S. alpinus complex.
Включение множества форм гольцов в S. alpinus complex нам представляется оправданным. Во-первых, это целесообразно на данном этапе познания с точки зрения практической систематики, т.к. в аналогичных ситуациях необходим определенный период накопления и осмысления материала. В противном случае система перестает быть системой, ибо гольцы дают возможность выделять новые таксоны чуть ли не в каждом озере. Во-вторых, в настоящее время экспериментально доказана обратимость изоляции у некоторых форм гольцов, возможность перехода одних форм в другие на протяжении жизни одного поколения, возможность быстрого проявления отклонений в значении ряда меристических признаков при вселении гольцов в новые водоемы [Frost, 1965; Roberts, 1971; Skreslet, 1973; Nordeng, Andersen, 1985; Nordeng, Hindar, 1985; Nordeng, Skurdal, 1985].
Теоретические аспекты возникновения различных жизненных форм в пределах одного вида в различных экологических условиях рассматриваются Б.М. Медниковым [1987]. Согласно его концепции, способность к существованию в качестве различных форм (дискретных адаптивных норм) наследуется при видообразовании от предковых видов и реализуется путем изменения экспрессии соответствующих генов, для которого факторы среды служат пусковым сигналом.
Говоря о гольце Сахалина, мы будем употреблять названия «мальма» и «голец» как синонимы. На Сахалине обитают проходная, ручьевая и речная формы гольца. Проходная форма распространена по отохоморскому побережью острова, от пролива Невельского на западном побережье до реки Очепуха на восточном; южнее устья Очепухи, в бассейнах озер Айруп, Тунайча, в реках залива Анива и юго-восточного Сахалина она отсутствует. Ручьевая форма распространена повсеместно в ручьях и верховьях рек, там, где они имеют характер ручьев. Речная форма встречена нами только в бассейне Тыми. Обитающую на Сахалине мальму А.Я. Таранец [1936] относил к южному подвиду S. malma krascheninnikovi, который от номинативного камчатского отличается более крупной чешуей, меньшим числом позвонков и чешуи в боковой линии. Ареал этого подвида простирается от устья Амура до залива Петра Великого, включает Сахалин, Южные Курилы и Хоккайдо. Нашими исследованиями [Гриценко, 1975] подтверждается видовое единство сахалинской мальмы и проходного гольца Камчатки, а также отношение их друг к другу в качестве подвидов. По используемым в анализе признакам различия между сахалинскими и камчатскими популяциями незначительно превышают различия между сахалинскими популяциями [Гриценко, 19706]. Исключение составляют различия в числе пилорических придатков, позвонков и чешуи в боковой линии. Поскольку последние два признака коррелированны, мы ограничимся рассмотрением числа позвонков и пилорических придатков. Как видно из рис. 60, число пилорических придатков подвержено нечетко выраженной клинальной изменчивости. У северных популяций оно выше, однако не всегда северные популяции отличаются от южных при попарном сравнении. Так, отсутствуют различия между чукотской мальмой и мальмой сахалинской реки Поронай. Хотя следует заметить, что географически более близкие камчатские и сахалинские популяции различаются на уровне, превышающем подвидовой (CD = 1,28). Подобное различие существует по числу позвонков. В этом случае любое парное сравнение дает величину CD, превышающую подвидовой уровень (рис. 61). Оба признака образуют уступы в клине, которым соответствует постоянство их значений на огромных участках ареала.
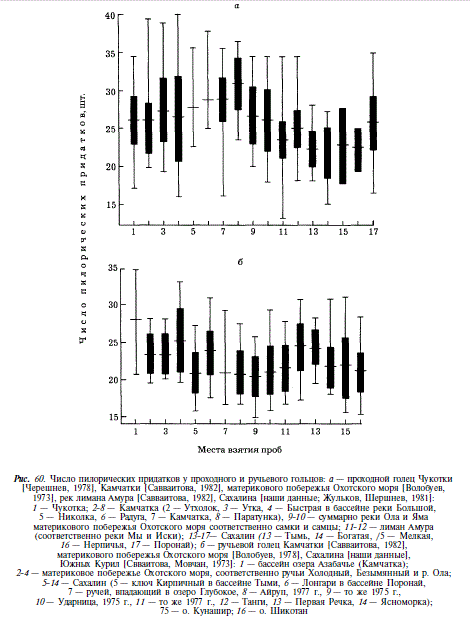
В качестве аргумента против использования числа позвонков как таксономического признака выдвигалась его изменчивость под воздействием различных температур в период раннего онтогенеза [Савваитова, Мовчан, 1973]. Однако едва ли можно назвать таксономический признак, который не изменялся бы под воздействием тех или иных факторов среды. В то же время, имеются многочисленные данные о высокой наследуемости числа позвонков [Кирпичников, 1979; Tanaka, 1969; McGregor, MacCrimmon, 1977; Leary et al., 1985; Beacham, Murray, 1986]. Кроме того, экспериментально установлено, что изменение числа позвонков на 1-2 происходит при различиях температуры воды в период инкубации в 5-7 °[Garside, 1966; Komoda, 1977]. В связи с этим становится ясно, что для получения различий в числе позвонков, подобных существующим между камчатскими и сахалинскими гольцами, нужны такие различия в температуре воды в период инкубации, какие не наблюдаются в природе. Однако даже если предположить, что различия в числе позвонков вызваны воздействием температуры воды, то следует говорить скорее о разных температурных предпочтениях камчатских и сахалинских гольцов, т.е. об экологических различиях. По мнению P.M. Викторовского [1975, 19786], реальность существования южного подвида мальмы подтверждается ее кариотипическими отличиями от гольцов Камчатки. Его данные, безусловно, не противоречат подвидовому подразделению, хотя их нельзя признать достаточными. Исходя из всего сказанного, мы считаем, что сахалинскую мальму следует рассматривать в отношении камчатской в качестве подвида, так, как это было предложено в свое время А.Я. Таранцом [1936], и назвать S. alpinus krascheninnikovi (Taranetz).
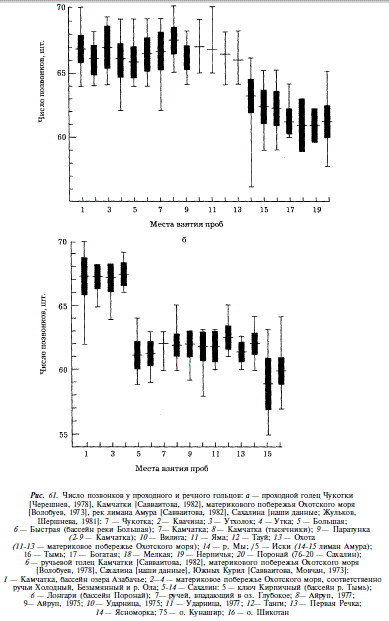
Ручьевым гольцом мы называем форму, описанную как S. malma curilus А.Я Таранцом [1936], который, учитывая значительную морфологическую обособленность, считал ее наследственно закрепленной. Размеры ручьевого гольца невелики. Минимальная длина половозрелой самки 10,8 см, максимальная — 25 см. От проходной формы ручьевая отличается относительно более длинной и высокой головой, более длинными челюстями, крупными глазами и длинными плавниками [Гриценко, 1975]. По числу жаберных тычинок, пилорических придатков и позвонков различий между проходной и ручьевой мальмами в целом по Сахалину не обнаружено. Однако в пределах бассейнов Тыми и Пороная у ручьевого гольца достоверно меньше жаберных тычинок, а в Тыми у него также меньше число позвонков (см. рис. 61, рис. 62). При сравнении сахалинских ручьевых гольцов с камчатскими проходными и ручьевыми гольцами, а также с гольцами из рек материкового побережья Охотского моря мы видим, что сахалинские ручьевые гольцы характеризуются, как и проходные, меньшим числом пилорических придатков и позвонков. При этом различия в числе позвонков более значительны, чем у проходных гольцов, что служит дополнительным основанием для выделения южного подвида мальмы.
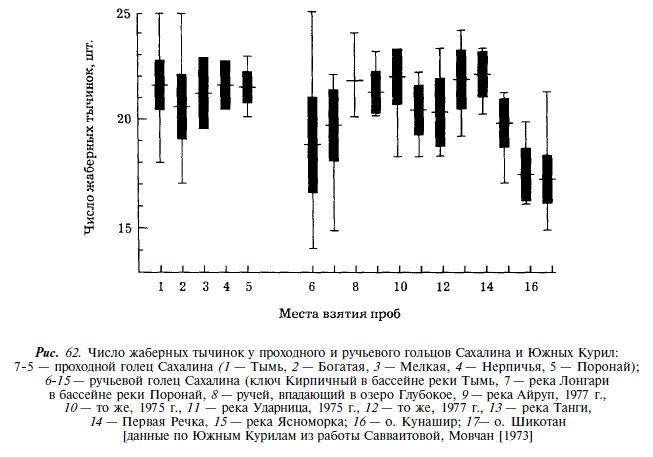
Южнокурильский ручьевой голец характеризуется наименьшим числом жаберных тычинок и позвонков, тем не менее он составляет по этим показателям единство с гольцами Сахалина, т.к. признаки эти гораздо меньше различны с таковыми у камчатских гольцов и гольцов материкового побережья Охотского моря. Если, следуя В.Л. Андрееву и др. [1978], выделять южнокурильского гольца в отдельный таксон, то он может рассматриваться лишь как natio по отношению к сахалинскому гольцу.
На Сахалине проходной и ручьевой гольцы репродуктивно обособлены. У них различаются сроки размножения, а в Тыми, кроме того, они населяют разные стации и крайне редко встречаются вместе [Гриценко, 1975].
Речной голец населяет верхнее и среднее течение Тыми и наиболее крупные притоки. По размерам он существенно превосходит ручьевого, достигая длины 42 см, массы 1,125 кг, внешне не отличается от проходного и в течение большей части года обитает совместно с проходным. Разделение половозрелых особей происходит лишь с конца июня до середины августа, когда проходной голец нагуливается в море. Учитывая это обстоятельство, мы для морфометрической характеристики речного гольца собирали только половозрелых особей в период с 15 до 30 июля, когда пребывание проходной формы в реке наименее вероятно. По морфологическим признакам речной голец несущественно отличается от проходного [Гриценко, 1975]. Форма эта выделяется нами провизорно. Весьма вероятно, что она является частью стада проходного гольца, не совершающей миграций в море.
Как следует из сказанного, голец на Сахалине характеризуется относительно простой популяционной структурой, что находится в соответствии с простотой гидрографической сети острова. Малые размеры рек и отсутствие озер, пригодных для обитания гольца, являются факторами, ограничивающими формообразование. Южнее, в Японии, там, где имеются горные озера, обитает и озерно-речная форма гольца [Kubo, 1967].






