Кижуч Oncorhynchus kisutch (Walbaum)
Кижуч распространен по азиатскому побережью от Чукотки до северной части острова Хоккайдо, по американскому побережью — от Аляски до Калифорнии [Берг, 1948; Линдберг, Легеза, 1965; Godfrey, 1965; Atkinson et al., 1967]. Сахалинские популяции обитают на периферии видового ареала. Распространение кижуча на Сахалине сходно с таковым осенней кеты: он встречается в реках восточного побережья и впадающих в залив Анива.
Наиболее численные популяции обитают на северо-востоке острова в реках Тымь, Набиль и ряде других, а также в бассейне реки Пороная. В годы депрессии численность стад отдельных рек составляет несколько сотен особей, в годы подъема — несколько тысяч, а в крупной реке Тымь — несколько десятков тысяч. На юго-востоке и в заливе Анива кижуч заходит в реки единично и не каждый год. В бассейне Пороная кижуч воспроизводится преимущественно в притоках среднего и нижнего течения. В 1965-1966 гг. в притоках верхнего течения молодь его нам обнаружить не удалось. В конце 70-х — начале 80-х годов отдельные особи кижуча стали повсеместно отмечаться в реках южного Сахалина, там, где их не встречали в течение 10-15 лет. Нам известны случаи захода их в реку Ударница (бассейн озера Тунайча). В значительном количестве молодь кижуча стала встречаться в верхней части бассейна Пороная. Таким образом, наблюдалась своеобразная небольшая "волна жизни".
На большей части ареала кижуч представлен летней и осенней формами [Смирнов, 1960, 1975; Зорбиди, 1975], наряду с доминирующей проходной у него имеются жилые формы [Двинин, 1949; Куренков и др., 1982]. У сахалинского кижуча А.И. Смирнов [1960,1975] также выделяет осеннюю и зимнюю сезонные формы. Нам, однако, их существование на Сахалине не представляется очевидным. По срокам хода сахалинского кижуча следует отнести к осенней форме, — около 80 % производителей заходят на нерестилища с конца сентября до конца октября. В Тыми начало хода отмечается в конце августа — начале сентября, в сроки, характерные для осеннего кижуча.
В сходные сроки кижуч заходит в реки Нерпичья [Жульков, 1976] и Набиль. Жилые формы кижуча на Сахалине не обнаружены.
Возрастная структура нерестовых стад кижуча определяется соотношением количества лет, проведенных особями в пресных водах и в море. При предельном абсолютном возрасте 4+ число возможных возрастных групп кижуча достигает по всему ареалу 10 [Коновалов, Щербинин, 1973]. Повсеместно основу численности составляют две возрастные группы: 2.+ и 3 2+ [Грибанов, 1948; Коновалов, Щербинин, 1973; Зорбиди, 1975; Godfrey, 1965; Godfrey et al., 1975]. Эта же особенность свойственна сахалинскому кижучу (табл. 29). Однако от популяций из других районов сахалинского кижуча отличает упрощенность возрастной структуры: отсутствие редких возрастных групп, а также наличие в крупнейшем нерестовом стаде (тымском) в течение нескольких лет лишь одной возрастной группы. Каюрки — особи, проведшие в море несколько месяцев и созревшие в год ската,
встречаются на Сахалине исключительно редко. В целом, созревание сахалинского кижуча происходит в более позднем возрасте, чем в большинстве районов ареала [Зорбиди, 1975; Волобуев, Рогатных, 1982; Godfrey, 1965].
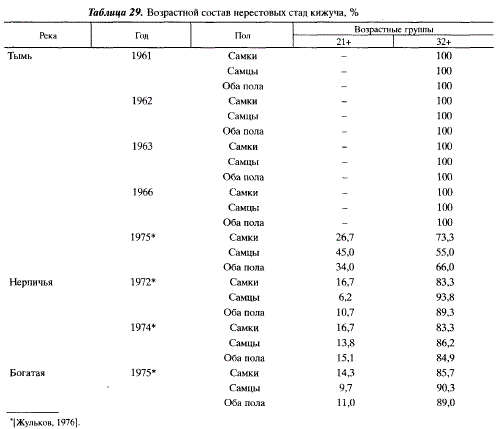
Длина и масса кижуча из разных районов обитания существенно различаются, а в пределах районов отмечаются значительные годовые различия. Камчатские популяции отличаются сравнительно небольшими размерами от популяций сахалинских и материкового побережья Охотского моря [Зорбиди, 1975; Волобуев, Рогатных, 1982а]. В пределах Сахалина наиболее крупный кижуч обитает в Тыми, наиболее мелкий — в Найбе (табл. 30). Самцы обычно бывают крупнее самок, при этом длина и масса первых варьирует в более широких пределах. Так, в Тыми за все годы наблюдения длина самцов колебалась от 59 до 83 см, масса тела — от 2 до 7 кг. У самок эти показатели равнялись соответственно 6178 см и 2,15-6,00 кг.
Индивидуальная абсолютная плодовитость кижуча колеблется в широких пределах: от 1760 до 9010 икринок, среднегодовая плодовитость отдельных стад — от 4320 до 5370 икринок. Она мало отличается от плодовитости кижуча материкового побережья Охотского моря [Волобуев, Рогатных, 19826] и Камчатки [Грибанов, 1948; Зорбиди, 19706]. В то же время, плодовитость азиатских популяций на 1/3 превышает плодовитость американских [Foerster, Pritchard, 1936; Wickett, 1951]. Последнее обтоятельство говорит о том, что американские стада находятся под меньшим давлением отбора, воспроизводятся в более благоприятных условиях, что находит выражение в большей численности кижуча в Америке, чем в Азии.
Соотношение полов в нерестовых стадах сахалинского кижуча приблизительно 1:1 при некотором преобладании самок [Гриценко, 1973; Жульков, 1976].
Сахалинскому кижучу свойственен наиболее ранний нерест по сравнению с кижучем Камчатки и Северной Америки. Около 80-90 % особей в Тыми нерестятся в период со второй половины октября до конца второй декады ноября. Ко второй половине декабря нерест практически заканчивается. Производители в начале хода поднимаются в верхние горные притоки, основная же масса нерестится во внутренней дельте Тыми и на участке основного русла выше поселка Красная Тымь. Нерест кижуча протекает преимущественно после спада осеннего паводка. Среднесуточная температура воды от начала к концу нереста понижается от 9 до 1 ° С. Производители устраивают бугры на выходах грунтовых вод, разрывая иногда при этом ранее устроенные бугры кеты [Гриценко, 1973]. Возможность конкурентных отношений из-за нерестилищ между кижучем и другими лососями предполагал еще И.И. Кузнецов [1928]. В настоящее время из-за низкой численности кижуча на Сахалине подобная конкуренция невелика. Нами было вскрыто 11 бугров кижуча в р. Тымь. Количество эмбрионов, обнаруженное в них, колебалось от 5 до 1441 шт., составляя в среднем 642 шт., или 12,8 % от средней за годы наблюдений плодовитости, что гораздо ниже соответствующего показателя у горбуши и кеты. Длина вышедших из грунта (или вымытых паводком) особей колеблется от 27 до 38 мм (составляя в среднем 31 мм), масса — от 150 до 550 мг (в среднем 260), у 57 % рыбок имелся остаток желточного мешка, масса которого составляла от 1,3 до 20,9 % (в среднем 9,4%) массы тела.
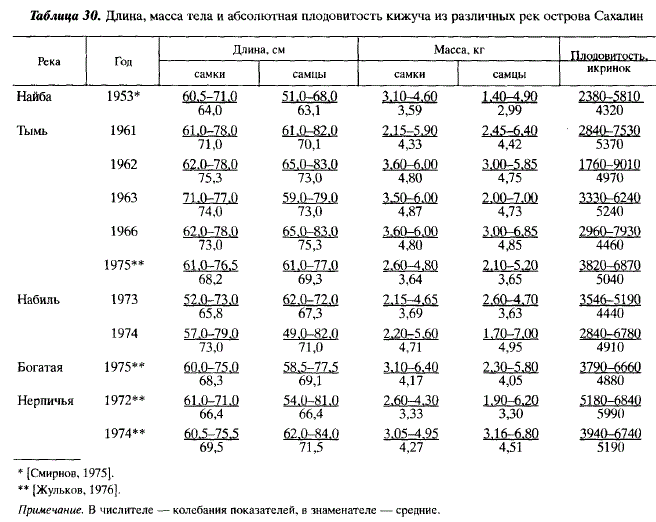
В первые дни и недели после выхода из грунта мальки кижуча расселяются по водоему. Часть из них разносится течением, часть расселяется в результате агрессивного территориального поведения, при котором крупные особи вытесняют мелких [Chapman, 1962]. К июлю мальки осваивают основные стации, обитание в которых свойственно кижучу: мелководные участки в районе нерестилищ, приглубые участки, имеющие древесные завалы, старицы и отшнуровавшиеся лужи на прибрежных косах. Часть мальков мигрирует вверх по течению, что неоднократно отмечалось при экспериментальных исследованиях американских популяций [Tripp, McCart, 1983]. В отличие от других лососей молодь кижуча часто нагуливается в местах с полным отсутствием течения и относительно высокой температурой воды (18-20 °С). В ручьи сеголетки кижуча заходят в относительно небольшом количестве, молодь проникает в них преимущественно на втором году жизни. Часть 2-леток-пестряток кижуча нагуливается на значительном удалении от нерестилищ на плесах среднего течения Тыми, где она становится равнинной рекой. В этой стации из лососевых рыб обитает, кроме молоди кижуча, только молодь сахалинского тайменя. Особенности распределения молоди кижуча по разным стациям наряду с иерархическими агрессивными внутривидовыми отношениями определяются также конкурентными отношениями с молодью других видов лососей [Hartman, 1965]. О характере распределения молоди кижуча в бассейне Тыми дает представление табл. 31.
Серебрянки кижуча начинают встречаться в верховьях рек в большом количестве с конца мая, однако скат происходит гораздо позже. В 1963 и 1965 гг. в среднем течении Тыми покатная миграция отмечалась со II декады июля по I декаду августа. В лимане рек Богатая и Нерпичья покатная молодь кижуча появлялась в конце июня, массовый скат отмечался во второй половине июля, заканчивался он в начале августа (рис. 51). Отдельные особи в лимане реки Нерпичья наблюдались до конца сентября [Чуриков, 19756; Жульков, 1976]. В реках Калифорнии скат происходит в более ранние сроки: с конца марта по начало июня с пиком, приходящимся на май [Shapovalov, Taft, 1954]. Однако как на северном Сахалине, так и в Калифорнии он начинается на спаде волны весеннего паводка, основная же масса молоди скатывается при уровне, близком к меженному. Среднесуточная температура воды в период ската колебалась в реке Богатая от 7,5 до 14,9 °С, во время массового ската — от 9 до 12 °С. В лимане реки она была на 0,5-1,0 °С выше. Температура в море в это время была равна в среднем 4,5 °С и колебалась от 1,6 до 7,5 °С. Перепад температуры воды при выходе из реки, следовательно, составлял от 7,5 до 10,5 °С.
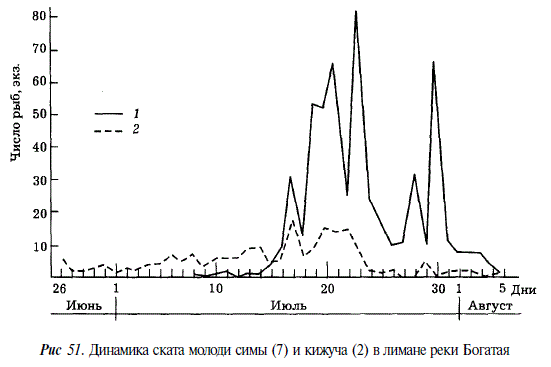
В качестве одного из факторов, определяющих выживаемость молоди кижуча при попадании ее в море, можно рассматривать резкий перепад температур речной и морской воды. При этом, очевидно, значение имеет не только величина перепада, но также и абсолютная величина температуры морской воды. Так, наблюдавшийся 16 июля перепад температуры от 11,0 °С в реке до 1,6 °С в море мог сказаться отрицательно на состоянии молоди симы и кижуча. Подобного рода перепады температуры должны особенно сильно влиять на молодь, скатывающуюся из небольших рек, сток которых не может существенно влиять на термический режим предустьевых участков моря.
Покатники кижуча в лимане Богатой были представлены двумя возрастными группами (1+ и 2+), из которых последняя резко преобладала (88,3 % самцов и самок). Среди самцов 3-леток было больше, чем среди самок (91,7 % против 85,6 %). На протяжении ската возрастной состав кижуча существенно не изменялся (табл. 32).
Также постоянны были и размеры кижуча. Самцы и самки по размерам почти не отличались (табл. 33).
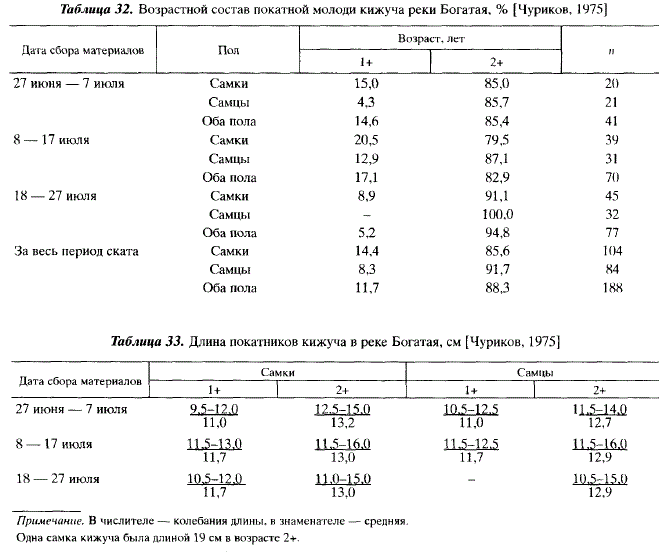
Масса покатников кижуча колебалась от 15 до 34 г (в среднем 25,1 г). Самцы и самки по массе не различались. Количество самок увеличивалось от начала ската к концу. С 27 июня по 7 июля самки составляли среди покатников 48,8 %, с 8 по 17 июля — 55,7 %, с 18 по 27 июля — 58,4 %. В целом за период ската соотношение самцов и самок было приблизительно 1 : 1 при некотором преобладании самок (55,3 %).
Аналогичная картина наблюдалась А.И. Жульковым в лимане реки Нерпичья, впадающей в Охотское море в нескольких десятках километров южнее Богатой. В реке Нерпичья также основное количество молоди кижуча (75,5 %) скатывалось в возрасте 2+. Размеры покатников были такими же, как в Богатой.
Сравнительно поздний скат молоди и ранний заход производителей кижуча на Сахалине, видимо, являются ведущими причинами практического отсутствия в нерестовом стаде каюрок (рыб в возрасте 2+), обычных в популяциях Северной Америки. В Калифорнии разрыв между временем массового ската молоди и захода производителей составляет около полугода, а на Сахалине — около 2 мес. Естественно, что за столь короткое время рыба не успевает вырасти и созреть.
В реках материкового побережья Охотского моря каюрки составляют 0,3-0,5 % нерестовых стад кижуча. Встречаются они в середине или конце хода [Волобуев, Рогатных, 1982б].
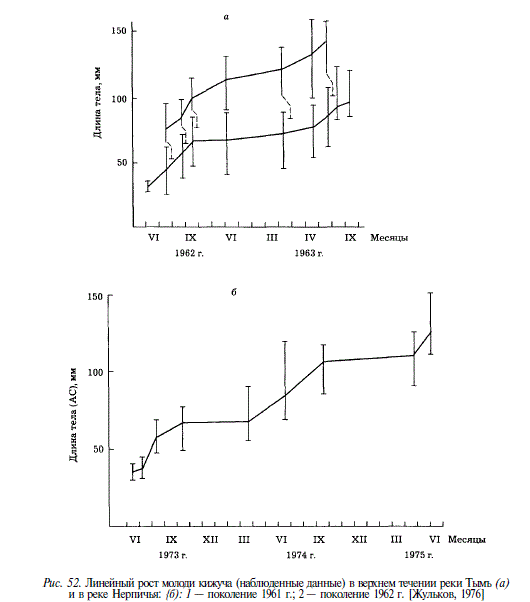
Рост молоди кижуча в течение пресноводного периода неодинаков в разные сезоны. У сеголеток он бывает наибольшим в период с мая по сентябрь, в дальнейшем — с сентября по апрель май — отмечается его резкое замедление или прекращение, с апреля по сентябрь вновь отмечается ускорение роста. У 2-леток кижуча в Тыми зимой 1962-1963 гг. рост замедлялся, но не прекращался. А.И. Жульков [1976] в аналогичной ситуации отмечал прекращение роста молоди кижуча в Нерпичьей. В предпокатный период, с мая по июнь, у 3-леток наблюдается очередное значительное увеличение длины (рис. 52). Зимнее замедление роста у особей американских популяций кижуча, обитающих в южной части ареала, не столь продолжительно, как у азиатских, и длится 1,52,5 мес, с ноября по начало февраля [Chapman, 1962].
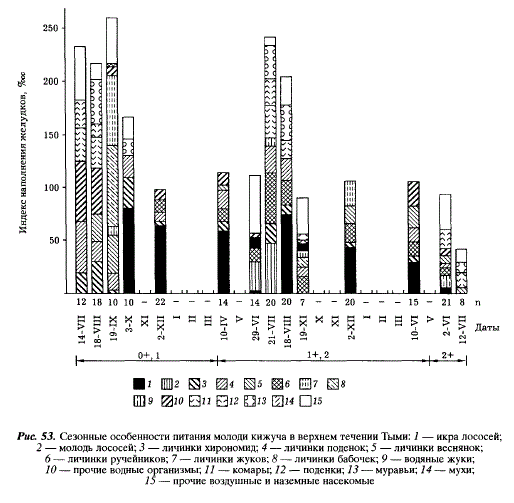
Исследуя питание молоди кижуча в пресных водах, мы рассматривали его сезонные, биотопические и суточные особенности. В Тыми наибольшие индексы наполнения желудков отмечаются у молоди всех возрастов летом и в начале осени, зимой происходит их снижение в 2-3 раза (рис. 53). Так же невелики они у покатной молоди в июле. С июня по сентябрь в пище всех возрастных групп доминируют воздушные и наземные насекомые, составляя по массе 40-50 % пищевого комка. Среди них преобладают имаго двукрылых, поденок, муравьев, среди донных организмов — личинки поденок, веснянок, ручейников, бабочек, водяных жуков, хирономид. Значение личинок веснянок и поденок остается относительно постоянным на всем протяжении пресноводного периода. Важным пищевым компонентом является икра тихоокеанских лососей, которую молодь начинает потреблять уже на первом году жизни. С осени до весны корм этот доминирует в желудках кижуча. Таким образом, годовые особенности питания молоди кижуча на Сахалине определяются ходом сезонных явлений в населяемых ею водоемах: преобладанием тех или иных стадий насекомых и нерестом лососей. Наши результаты совпадают с полученными ранее на Камчатке [Зорбиди, 1970а]. Однако в отличие от камчатского кижуча, молодь которого потребляет преимущественно хирономид, в пище сахалинского кижуча хирономиды играют второстепенную роль.
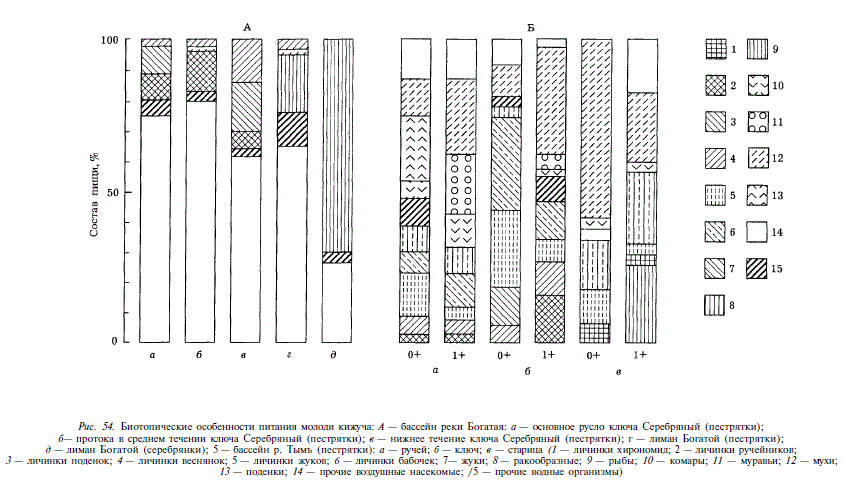
Биотипические особенности питания молоди кижуча исследовались в различных стациях верхнего течения Тыми в августе (ручей, ключ, старица), а также в бассейне Богатой в июле (среднее и нижнее течение ключа Серебряный, лиман) (рис. 54). Почти во всех случаях в рационе кижуча доминировали воздушные беспозвоночные, видовой состав которых, однако, различался в различных стациях. Среди донных беспозвоночных, потребленных кижучем в проточных водах, преобладали личинки веснянок, поденок и ручейников, в стоячих и слабопроточных водах (старица, ключ) — личинки бабочек и жуков, а также имаго жуков. В лимане Богатой в пище молоди кижуча появляются ракообразные (преимущественно бокоплавы), которые в рационе серебрянок резко доминируют. Обращает на себя внимание крайне редкая встречаемость в пище молоди кижуча рыб.
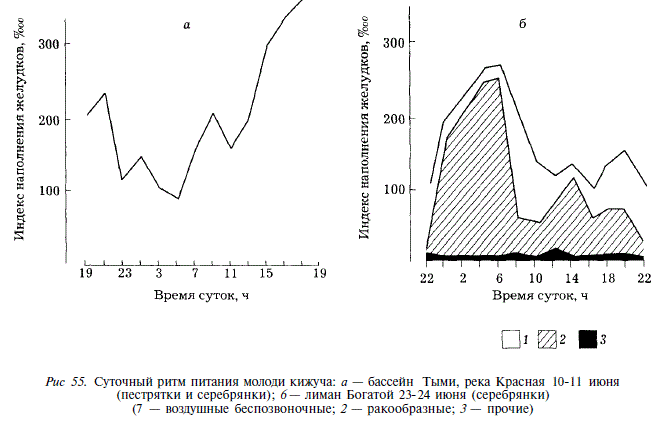
В суточной активности питания молоди кижуча в верховьях Тыми (в реке Красная) в начале июня отмечается два подъема: с 5 до 9 и с 11 до 17 ч. С 17 до 19 ч наполненность желудков остается высокой, с 19 до 5 ч она постепенно снижается до минимума [Гриценко, Ардавичус, 1967] (рис. 55). В лимане Богатой в суточной пищевой активности серебрянок отмечаются три подъема. Наибольшая активность свойственна молоди в ночные часы, в это время в пище доминируют ракообразные, поднимающиеся в толщу воды, с рассветом увеличивается доля воздушных насекомых. Незначительное повышение активности отмечается в 12-14 ч. В вечерние часы наступает новое повышение активности, при котором молодь активно охотится за летающими над водой насекомыми.
Суточные особенности питания животных определяются чередованием преобладания пищевого и оборонительного рефлексов [Гирса, 1962; Мантейфель, 1980]. Поскольку в реках молодь кижуча крайне редко встречалась в желудках хищных рыб, что свидетельствует о практическом отсутствии у нее врагов, суточные особенности ее питания определяются в основном наличием и доступностью корма, но не оборонительной активностью. Сезонные и суточные особенности питания молоди кижуча во многом определяются характером дрифта беспозвоночных [Hugt, 1975; Чебанова, 1983]. Особое место принадлежит при этом поверхностному дрифту. Естественно, что в стоячих водоемах, а также в зимнее время, когда кижуч концентрируется на участках с замедленным течением, он в большей степени становится бентофагом, чем в проточных водах и в летние месяцы. Таким образом, основные особенности питания молоди кижуча остаются довольно постоянными на всем разнообразии населяемых им стаций. Даже резкая физиологическая перестройка, связанная со смолтификацией, и выход в предустьевые участки, отличающиеся по гидрологическим особенностям от условий обитания в реках, не вызывали заметных изменений в характере питания: по-прежнему основным кормом оставались воздушные насекомые.
Численность производителей кижуча в верхнем течении Тыми колебалась с 1961 по 1986 г. от 1 до 40 тыс. экз., обычно не превышая 10 тыс. экз. Отмечалось два подъема его численности: в 1961 и в 1983-1986 гг. В других реках северо-восточного Сахалина численность кижуча измеряется сотнями, изредка — тысячами особей. В промысле сахалинский кижуч существенной роли не играет. Основным фактором, сдерживающим его численность, является вылов в реках. Производители добываются в большом количестве браконьерами, а молодь — многочисленными рыбаками-любителями, вылавливающими ее на удочку. При существующей обстановке едва ли приходится ожидать увеличения численности кижуча только за счет усиления охраны. Реальным может быть создание промысловых стад кижуча за счет искусственного разведения при условии, что молодь будет выпускаться на стадии смолта, и на реках, где будут расположены заводы, будет установлен заповедный режим.






