Экология молоди кеты в период ската
Первые мальки кеты в верховьях Тыми обычно выходят из бугров в последних числах марта — первых числах апреля. Появившиеся стайки мальков держатся в мелководных прогретых заливах, в ключах и лимнокренах. Обычно они находятся вблизи полуразложившихся трупов производителей кеты, около которых в изобилии держатся мелкие личинки веснянок, поденок и хирономид, служащие малькам пищей. К середине — концу апреля количество нагуливающихся мальков возрастает, они образуют скопления но несколько тысяч, и высокая плотность скоплений вынуждает их рассеяться по водоему. До начала мая ската как такового не наблюдается, перемещения молоди в пределах внутренней дельты Тыми легко отмечаются визуально. Поскольку значительная часть бугров кеты находится выше уреза воды, выход из них возможен лишь после подъема уровня.
Скат кеты в Тыми начинается обычно в первых числах мая, но иногда в конце I — начале II декады мая. В среднем скат в районе села Кировское длится 64 дня, в районе села Адо-Тымово — 86 сут., в реке Набиль — 48 сут. (табл. 26). Таким образом, намечается закономерность увеличения продолжительности ската кеты по мере увеличения размеров нерестового водоема и увеличения численности заходящих на нерест лососей. Продолжительность ската, по-видимому, связана также с продолжительностью нерестового хода. В Набили нерестовый ход намного короче, чем в Тыми. В верховьях Тыми 50 % покатной молоди успевает скатиться к 26 мая, в Набили — к 13 июня. Соответствующее запаздывание набильской молоди отмечается и в сроках ската 75 % молоди. Видимо, обильные выходы теплых грунтовых вод на нерестилищах кеты в Тыми способствуют более быстрому развитию ее молоди и более раннему скату. Ко времени ската 90 % молоди этот разрыв уменьшается, и полное завершение ската в верховье рек Тымь и Набиль наступает в одни и те же сроки. В среднем течении Тыми, у села Адо-Тымово, скат завершается почти на месяц позже — 5 августа. В продолжение последнего месяца скат бывает крайне малочисленным. Скатывающаяся в течение июля — начале августа молодь активно питается в реке. Экологически она соответствует серебристым пестрякам симы и кижуча.
В верховьях Тыми скат бывает наиболее интенсивным вначале и постепенно ослабевает к концу, в среднем ее течении — во время ската третьей четверти молоди. Последние 10 % молоди скатываются крайне медленно. В Набили наибольшая интенсивность ската наступает уже после того, как скатится 50 % молоди. И во всех трех пунктах мы наблюдаем ослабленную интенсивность ската последних 10 % молоди (табл. 27).
А.А. Чуриков [19756] описал аналогичную задержку небольшого количества молоди кеты в эстуарии реки Мелкая, где она нагуливалась в течение второй половины июля. Задержка небольшого количества молоди в верховьях Тыми до последних чисел августа — начала сентября отмечалась и нами [Воловик, Гриценко, 1968]. Можно предположить, что такая задержка свойственна потомству производителей наиболее поздних сроков нереста, единично встречающихся в реке вплоть до марта. В конце сентября молодь кеты перестает встречаться в Тыми, видимо, скатываясь во время осенних паводков.
Анализируя совмещенные графики суточного ската кеты и горбуши, нельзя не отметить их большого сходства. В качестве наглядных примеров можно привести скат в верховьях Тыми в 1977 г. (см. рис. 35) и в ключе Холодный в 1976 г. (см. рис. 36). Менее отчетливо такое сходство прослеживается и в другие годы. Это обстоятельство позволяет предположить, что ряд факторов оказывает сходное действие на покатную миграцию молоди горбуши и кеты. В частности, сходным образом изменяется скат обоих видов в связи с изменениями температуры воды. Частые и резкие изменения температуры и уровня воды обусловливают колебания интенсивности ската кеты подобно тому, как это происходит у горбуши. Такое сходство естественно и понятно, поскольку молодь этих видов мигрирует в море сеголетками приблизительно одной массы, имея подчас значительные остатки желточного мешка. Кроме того, в верховьях рек значительная часть молоди кеты скатывается, не питаясь, подобно горбуше.
Вместе с тем, нельзя не заметить, что скат молоди кеты в начальный период в большей мере, чем скат горбуши, определяется влиянием паводков. Большая продолжительность ската кеты в среднем течении Тыми, по сравнению с горбушей, объясняется тем, что первая задерживается в реке для нагула, что горбуше как виду не свойственно.
В Поронае завершение ската кеты так же растянуто во времени, как и в Тыми. Однако в Поронае и горбуше нередко свойственно такое завершение. Это, видимо, следует объяснить присутствием в скате потомства охотоморской осенней горбуши, которая отсутствует в Тыми.


Изменения суточной активности молоди кеты от начала к концу ската и от верховьев к низовьям рек напоминают таковые у горбуши. В верховьях рек в районе нерестилищ кете присущ ночной скат, продолжительность которого уменьшается по мере сокращения темного времени суток и увеличения прозрачности воды. Так, в I декаде мая суточный период ската колеблется в разные годы от 540 до 600 мин, в I декаде июля — от 240 до 300 мин. В отдельные годы в пик паводка незначительная часть молоди кеты в верховьях Тыми скатывается в светлое время суток. В среднем течении крупных рек в начальный период ската молодь мигрирует круглосуточно, а в конечный период ей свойственна четко выраженная ночная активность (рис. 48).
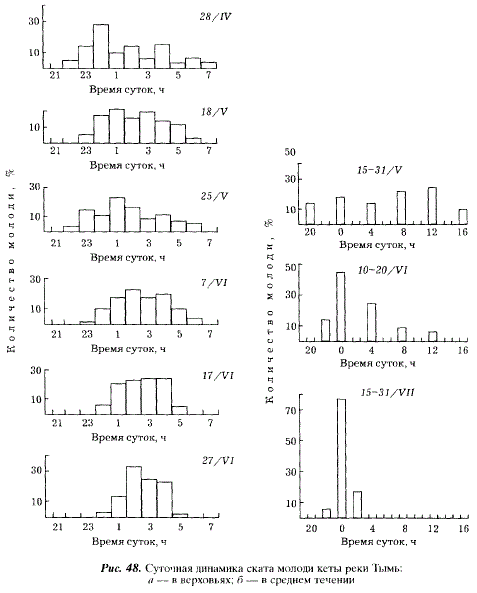
Подобно горбуше молодь кеты Тыми не задерживается на нагул в Ныйском заливе, а выносится из него мощными приливо-отливными течениями. При миграции через залив молодь не распространяется за пределы фарватеров. Выход в море кеты происходит в отлив или в начале прилива.
Существенно отличается от этого начало морского нагула у кеты юго-западного Сахалина, которая в течение полутора месяцев обитает в прибрежных мелководьях, служащих для нее своеобразным питомником. Хорошая прогреваемость, обильный корм и пониженная соленость этих участков облегчает молоди адаптацию к морским условиям [Шершнев, 1971]. Если на северо-восточном Сахалине выживаемость молоди кеты лимитируется низкими температурами воды в море, то на юго-западном лимитирующим фактором становятся высокие температуры. Соответственно, на северо-востоке между Кв и термикой моря существует, как мы писали выше, положительная корреляция, на югозападе — отрицательная [Шершнев, 1974].
Ю.С. Рослый [1975а], изучая посредством витального окрашивания рыб скорость миграции молоди кеты в низовьях Амура, установил, что она колеблется от 69,4 до 127,2 км/сут. Такие различия дали ему основание предположить, что одна часть молоди активно движется в потоке, а другая скатывается, медленно откочевывая к берегам для нагула. Его предположение подтверждается нашими наблюдениями. В среднем течении Тыми во второй половине июня — в июле, в светлое время суток, эти две группы визуально различаются очень хорошо. Одна из них мигрирует небольшими стайками в зоне высоких скоростей течения, другая также небольшими стайками по 5-50 экз. держится вблизи берегов, кормится и медленно сплывает вниз по течению, задерживаясь в очередном участке с замедленным течением. Состав стаек непостоянен: время от времени 1-2 рыбки отрываются от стайки и, проплыв несколько метров вниз по течению, присоединяются к другой. В принципе, состав стайки, состоящей из 50 особей, может полностью смениться за 4-5 ч. Скорость перемещения нагуливающихся стаек кеты в дневное время колеблется от 30 до 100 м/ч. С наступлением сумерек эта молодь отходит от берега и скатывается в участках русла с быстрым течением.
Ю.С. Рослый наблюдал подобное перемещение молоди кеты к берегам для нагула в нижнем течении Амура. В низовьях Амура молодь задерживается на нагул ночью и скатывается днем. Он связывает эту миграцию с суточным ходом изменения температуры воды. Отличия его наблюдений от наших, видимо, обусловлены различиями стаций. В низовьях крупных рек скат молоди приобретает новые черты по сравнению со скатом в верхнем и среднем течении.
У некоторой части скатывающейся молоди кеты имеются остатки желточного мешка, составляющие до 37 % от массы тела. Мы заметили, что количество молоди, скатывающейся с нерассосавшимся желточным мешком, различно как по годам, так и по периодам ската.
На протяжении сезона ската число мальков с большим содержанием желтка у молоди кеты увеличивается с подъемом и уменьшается с понижением уровня реки. В 1966 г. весенний паводок был слабее предыдущего, но количество молоди с большим содержанием желтка оказалось выше (рис. 49). В отдельные сутки оно составляло 40 %, в то время как в 1965 г. не превышало 23,3 %. При пересчете на всю скатывающуюся молодь разница оказалась еще значительнее. За период с 18 мая по 26 июня 1965 г. скатилось 9485 тыс. мальков и личинок кеты (или 47 % всего ската), причем личинок было 576 тыс. (6,1 %). С 19 мая по 29 июня 1966 г. скатилось 24231 тыс. экз. (76,9 % всего ската) и из них в стадии личинок — 4726 тыс. экз. (19,5 %). Эти различия объясняются условиями нереста родительского стада. В 1964 г. нерест прошел при условиях, близких к оптимальным. Эмбрионы успели закончить развитие к началу весеннего паводка. Размывание гнезд нанесло ущерб только немногочисленному потомству позднего нереста. В 1965 г. значительная часть гнезд раннего и массового нереста была уничтожена осенним паводком, поэтому в скате 1966 г. было больше молоди от позднего нереста, не успевшей достаточно развиться. Таким образом, осенние паводки, вызываемые тайфунами, приводят не только к уменьшению количества покатной молоди, но и к снижению ее жизнестойкости. Покатники, имеющие большой остаток желтка, едва ли могут считаться жизнестойкими. Они чаще, чем мальки, становятся добычей хищных рыб, составляя в частности существенную часть рациона мальмы. Для установления критерия жизнестойкости личинок мы провели два наблюдения. В первом из них определялась величина остатка желтка у молоди, активно вышедшей из грунта и нагуливающейся в районе нерестилищ. Среднее содержание желтка составляло у рыб 2,5 % массы тела, максимальное — 6,6 %. По нашим наблюдениям, особи с остатком желточного мешка в 6-8 % обладают такой же подвижностью и маневренностью, как и молодь, у которой желток полностью рассосался.
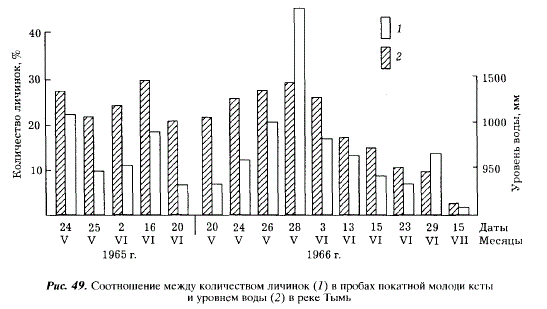
Во втором случае мы руководствовались тем обстоятельством, что молодь кеты в верховьях Тыми скатывается ночью, днем же она заходит в мелководные заливы, где есть обильный корм и куда нет доступа хищным рыбам. Молодь, заходящую днем в такие места, мы относим к активно скатывающейся и содержание желтка у нее принимаем за нормальное. У молоди, выловленной ночью на стрежне реки, среднее содержание желтка составило 6,5 %, максимальное — 15,9 %, у молоди, пойманной в 11-12 ч у берега, среднее содержание желтка было 4,3%, максимальное — 11,2 %. В связи с этим мы условно приняли за критическое содержание желтка, равное 10 % массы тела. Относительное количество рыб, имевших больше желтка, принимается за показатель, в какой-то мере характеризующий степень размыва нерестовых гнезд кеты весенним паводком.
Размеры и масса покатной молоди кеты закономерно изменяются от начала к концу ската (табл. 28). В первые дни ската, при еще низком уровне воды идет молодь, распределение длины которой характеризуется четко выраженной правой асимметрией и относительно большой вариабельностью (рис. 50). В дальнейшем, по мере подъема уровня, идет молодь меньших размеров. Распределение ее по длине в отдельных пробах отличается левой асимметрией за счет вымывания из грунта особей с желточным мешком. Вариабельность длины уменьшается. В процессе паводка происходит незначительное увеличение длины и массы мигрантов, однако размерный ряд остается узким. Размеры молоди, скатывающейся после прохождения паводка в период летнего падения уровня, резко увеличиваются. Распределение молоди по длине вновь характеризуется правой асимметрией, ряд становится более широким. В самом конце ската размеры молоди вновь несколько уменьшаются, хотя в целом характеристика размерного ряда остается прежней. Подобное уменьшение наблюдалось не каждый год, но приблизительно в трети лет в период с 1960 по 1981 г.

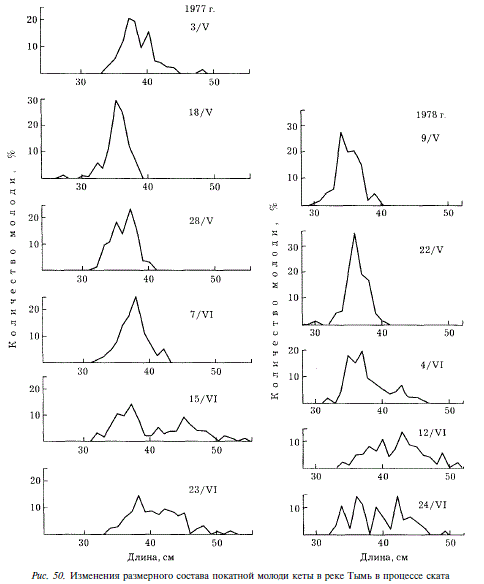
Аналогичная ситуация наблюдалась Ю.С. Рослым [1967] в бассейне Амура. Этот автор также выделяет среди покатной молоди кеты две группы: молодь с прогонистым челом пятнистой окраски и с неполностью рассосавшимся желточным мешком; высокотелую молодь серебристой окраски, упитанную и с рассосавшимся желточным мешком. Мальки второй группы задерживаются в реке для нагула и откармливания в прогреваемых заливах. Их количество среди мигрантов увеличивается к концу ската.
Приведенные выше данные но размерному составу покатной молоди и срокам ската, в частности, растянутость во времени ската последних 10 % мигрантов, дают основание говорить об определенной разнокачественности покатной молоди кеты, о существовании двух групп мигрантов и двух компонентов ската. Первый компонент включает особей длиной от 27 до 40 мм, скатывающихся преимущественно в период паводка, мало связанных с нагулом в реке, торопящихся покинуть ее, подобно молоди горбуши, для нагула в море. Второй компонент включает особей длиной от 40 до 54 мм. Эта молодь активно питается и растет в реке, увеличивая первоначальную массу в 2-3 раза. Период нагула ее в пресных водах составляет 1-2 мес. Скат этой крупной молоди наблюдается до или после прохождения паводка. В паводок она держится на участках с замедленным течением, в ключах, лимнокренах и на разливах, где активно питается. Одной из вероятных причин, обусловливающих скат этой молоди, можно считать плотность скоплений в местах нагула. В начальный период ската она увеличивается за счет выхода из грунта все большего количества молоди, в конечный период — за счет падения уровня и сокращения нагульных площадей. Эта категория мигрантов имеет много сходства с покатной молодью кижуча, симы, нерки и чавычи. Первый компонент ската кеты можно условно назвать горбушеподобным, второй — кижучеподобным.
В онтогенетическом плане причину возникновения двух компонентов, видимо, следует искать в физиологической разнокачественности молоди кеты, в разной готовности ее к обитанию в морской воде, в разной скорости протекания процессов смолтификации. В плане эволюционном смысл этой разнокачественности в большей защищенности вида во времени за счет растянутости сроков ската и разнообразия размеров покатников. Как считает Э.Л. Бакштанский [1968], введший понятия "защищенность во времени" и "защищенность в пространстве", одним из основных элементов жизненной стратегии относительно малочисленных видов лососей, имеющих незначительные по площади нерестилища, является растянутость во времени основных жизненных стадий, например, сроков ската и возраста смолтификации, сроков созревания производителей, гарантирующая попадание хотя бы части особей популяции в благоприятные условия обитания.






