СЕЗОННАЯ ХАРАКТЕРИСТИКА ГОНАДОГЕНЕЗА ТРЕПАНГА ДАЛЬНЕВОСТОЧНОГО APOSTICHOPUS JAPONICUS SELENKA
Сезонные изменения развития гонад дальневосточного трепанга хорошо изучены (Низовская, 1971; Касьянов и др., 1980; Касьянов, 1989; Технология ..., 2001*). Танака (Tanaka, 1958), описывая репродуктивный цикл трепанга, обитающего у берегов южного Хоккайдо, выделил в нем стадии покоя, восстановления, роста, созревания и вымета. Л.В. Низовская (1971) разделила репродуктивный цикл самок трепанга из зал. Петра Великого на 4 стадии. По ее мнению, посленерестовая стадия непродолжительна, восстановительная стадия заканчивается в ноябре, стадия медленного роста захватывает зимние и первые весенние месяцы, начавшаяся в апреле стадия быстрого роста приводит гонаду в преднерестовое состояние. В.Л. Касьянов с соавторами (Касьянов и др., 1980; Касьянов, 1989) выделили в половом цикле трепанга три стадии: с конца июля и до середины августа — нерест, с середины августа и по октябрь — стадия восстановления, с октября по июль — стадия накопления и дифференциация гамет. Китайские исследователи разделили половое развитие трепанга на 5 периодов: пауза, восстановление, созревание, зрелость и нерест*. Относительно продолжительности стадии покоя единого мнения нет. По данным Л.В. Низовской (1971) и В.Л. Касьянова с соавторами (Касьянов и др., 1980; Касьянов, 1989), она занимает немногим более месяца, тогда как Танака (1958) указывает срок до трех месяцев. Китайские исследователи, описывая стадии развития гонады трепанга, считают, что продолжительность этой стадии составляет 4 мес*.
Цель настоящей работы — гистофизиологическое исследование гонад самок и самцов трепанга дальневосточного Apostichopus japonicus в различные сезоны года, анализ динамики клеточного состава половых желез и установление сроков нереста и продолжительности стадии покоя.
Материалы и методы
Исследовались гонады самок и самцов трепанга на полигонах ТИНРО-центра (бухты Киевка, Соколовская, Мелководная, Средняя и на акватории от мыса Гембачева до мыса Де-Ливрона) с мая по октябрь 2011 г. У трепанга определяли общую массу, массу кожно-мускульного мешка (КММ) и массу половых желез с точностью до 0,1 г. Гонадный индекс (ГИ) определяли как отношение массы гонады к КММ, выраженное в процентах. Пол животных и состояние гонады устанавливали визуально, а также на временных и постоянных гистологических препаратах. Гонады фиксировали в 70 %-ном спирте и растворе Буэна, проводили через Isoprep (производство БиоВитрум) и заливали в Histomix (производство БиоВитрум) согласно протоколу проводки (Микодина и др., 2009).
Полученные на санном микротоме срезы толщиной 5-6 мкм окрашивали гематоксилином Эрлиха с докраской эозином и заключали в синтетическую монтирующую среду Bio Mount (производство БиоВитрум). Готовые препараты исследовались в проходящем свете при различном увеличении (100, 400) на микроскопе Olympus model BHT-2 и фотографировались с помощью цифровой камеры Olympus. Стадии развития гонад определяли по наличию доминирующих репродуктивных клеток согласно существующим классификациям (Низовская, 1971). Кроме того, с помощью винтового окуляра-микрометра МОВ-1-15х измеряли диаметр половых трубочек (в миллиметрах), размер ооцитов (в микрометрах) и их количество в половых трубочках. Для гистологического анализа было взято 89 особей.
Технология разведения трепанга и морского ежа : пер. с кит. Циндао: Изд-во Рыбохоз. ун-та, 2001. 157 с.
Результаты и их обсуждение
Во время исследования соотношение полов у трепанга было равным во всех бухтах.
Трепанг имеет ветвистую непарную гонаду с половым протоком, проходящим по спинному мезентерию кпереди и открывающимся у основания щупалец. Система ветвящихся трубочек покрыта целомическим эпителием, который подстилают поперечные и продольные мышечные волокна. Глубже лежат тонкая соединительнотканная прослойка и базальная мембрана с примыкающими к ней половыми клетками. Летом пол особи можно установить по цвету гонады. У самцов она беловатого или бело-зеленого цвета, у самок — розоватого или ярко-оранжевого. Размер половой железы варьирует в зависимости от репродуктивного состояния особи, достигая 21 см в длину (Брегман, 1971). Возраст, в котором трепанг достигает половозрелости, трудно указать точно в связи с отсутствием у него регистрирующих структур и со значительным индивидуальным варьированием темпов роста и развития в зависимости от географического района, условий места обитания, сроков оседания личинок и других факторов (Бирюлина, Козлов, 1971; Брегман, 1971; Гаврилова, 2013). Считается, что к 2-3 годам особи могут приступать к нересту (Левин, 2000).
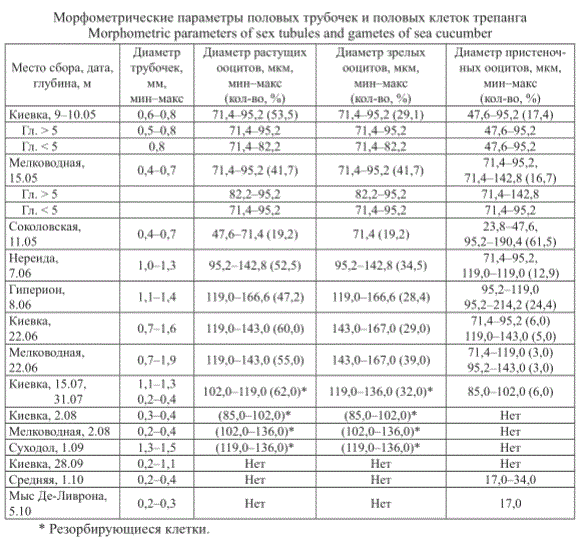
В исследованный период (с мая по октябрь) морфометрические параметры половых трубочек и гамет изменялись (см. таблицу).
Морфометрические параметры половых трубочек и половых клеток трепанга Morphometric parameters of sex tubules and gametes of sea cucumber
Диаметр Диаметр растущих Диаметр зрелых Диаметр пристеноч-
Место сбора, дата, трубочек, ооцитов, мкм, ооцитов, мкм, ных ооцитов, мкм,
* Резорбирующиеся клетки.
В первой и второй декадах мая у самцов и самок трепанга из бухт Киевка и Мелководная в гонадах идут активные гаметогенетические процессы (рис. 1 и 2), у самцов отмечается складчатость внутренней поверхности половых трубочек и процессы пролиферации гониев. В трубочках присутствуют сперматогонии и сперматоциты. У самок половые трубочки заполнены растущими ооцитами (рис. 1, 2).
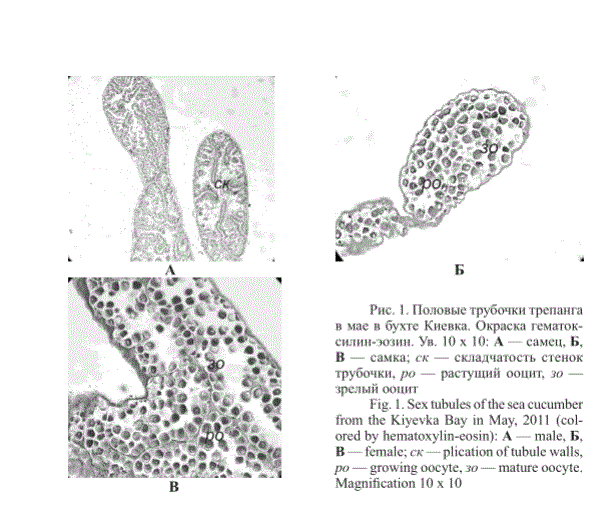
Рис. 1. Половые трубочки трепанга в мае в бухте Киевка. Окраска гематоксилин-эозин. Ув. 10 х 10: А — самец, Б, В — самка; ск — складчатость стенок трубочки, ро — растущий ооцит, зо — зрелый ооцит
В июне у большинства самок трепанга из бухты Киевка в половых трубочках находились растущие (на стадии протоплазматического и трофоплазматического роста) и зрелые ооциты. Редко встречались особи с половыми трубочками, в которых только начинался процесс пролиферации (рис. 3).
В первой половине июля у самок из бухты Киевка встречаются трубочки, заполненные растущими и зрелыми ооцитами, а также частично опустошенные (1/2 часть трубочки заполнена свободно лежащими ооцитами, а другая половина — пустая). Стенка половой трубочки в это время предельно истончена. В конце июля в половых трубочках самцов преобладают спермии, расположенные в центральной части трубочки, и сперматоциты II порядка (рис. 4).
В августе у большинства самок и самцов из исследованных бухт в полости тела были пустые половые трубочки с формирующейся складчатостью стенок (рис. 5). Встречались также особи, у которых находились трубочки с единичными невы-метанными зрелыми ооцитами и трубочки с резорбирующимися ооцитами (рис. 5). Как правило, после нереста у трепанга происходит резорбция невыметанных гамет и распад половых трубочек благодаря активности фагоцитирующих клеток. Известно, что при резорбции невыметанных половых клеток полость гонады очищается от гамет предыдущей популяции, тем самым освобождая пространство для прохождения нового гаметогенетического цикла (Касьянов и др., 1980; Касьянов, 1989). Кроме того, для следующей генерации половых клеток используются питательные вещества, накопленные в резорбируемых гаметах и составляющие дополнительный источник вещества и энергии для развивающихся ооцитов. Продукты резорбции гамет могут быть источником синтеза гормональных веществ, вырабатываемых гонадой. Дегенерация и резорбция гамет под влиянием эндогенных и экзогенных факторов нейросекреторной системы в свою очередь являются регулятором гамето-генетических событий. Без резорбции и дегенерации невыметанных зрелых гамет не начинается (или сдерживается) пролиферация половых клеток следующей генерации (Касьянов, 1989).
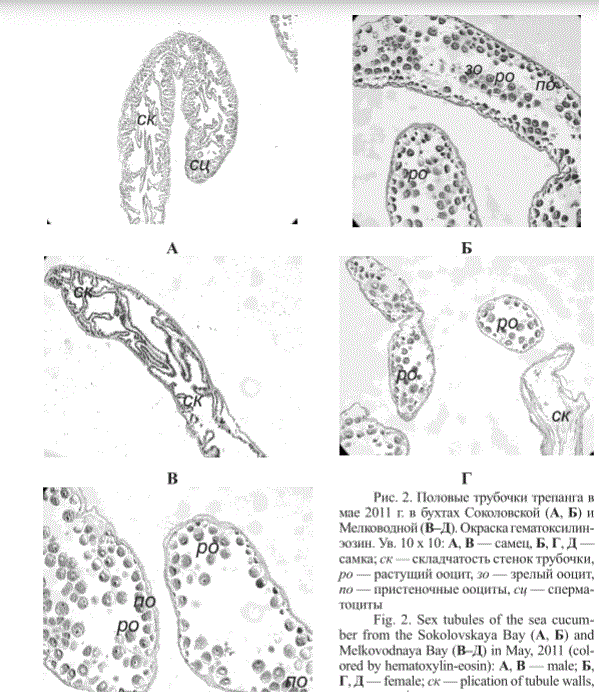
Рис. 2. Половые трубочки трепанга в мае 2011 г. в бухтах Соколовской (А, Б) и Мелководной (В-Д). Окраска гематоксилин-эозин. Ув. 10 х 10: А, В — самец, Б, Г, Д — самка; ск — складчатость стенок трубочки, ро — растущий ооцит, зо — зрелый ооцит, по — пристеночные ооциты, сц — сперма-тоциты
Fig. 2. Sex tubules of the sea cucumber from the Sokolovskaya Bay (A, Б) and Melkovodnaya Bay (B-Д) in May, 2011 (colored by hematoxylin-eosin): A, В — male; Б, Г, Д — female; ск—plication of tubule walls, ро — growing oocyte, зо — mature oocyte, по — wall oocytes, сц — spermatocyte. Magnification 10 х 10
В сентябре в бухте Киевка встречались единичные особи с половыми трубочками, заполненными резорбирующимися половыми клетками (рис. 6, А, Б). Большинство имели половые трубочки с толстыми стенками, образованными слоем цилиндрического эпителия толщиной от 17,0 до 68,0 мкм (рис. 6, В, Г). Во всех трубочках присутствовала складчатость внутренней поверхности стенок.
В первой декаде октября в бухте Средней у самок трепанга на поперечных срезах трубочек заметна складчатость их стенок (рис. 7, А). Вдоль внутренней поверхности лежат оогонии и растущие ооциты размером 17,0-34,0 мкм в количестве 35-158 шт. (в зависимости от размера трубочки) (рис. 7, Б-Е). У самцов, как правило, одна третья часть половой трубочки пустая, одна треть — со складчатостью стенок и одна треть — со скоплением герминативного эпителия, дающего начало новой ге-
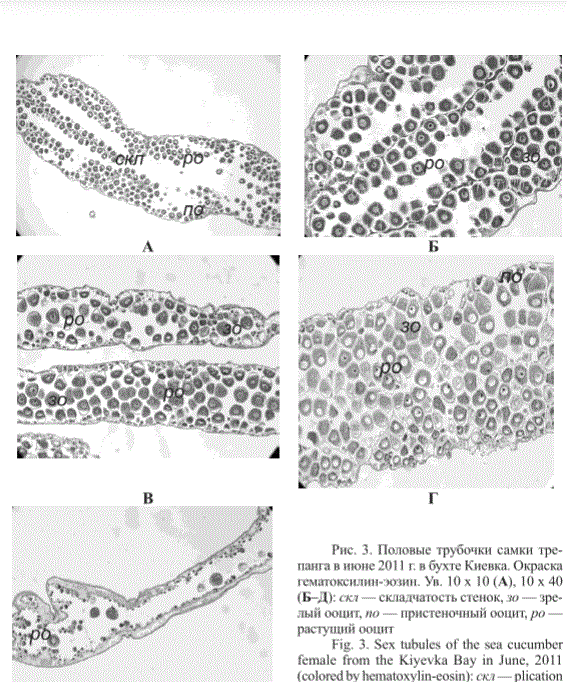
Рис. 3. Половые трубочки самки трепанга в июне 2011 г. в бухте Киевка. Окраска гематоксилин-эозин. Ув. 10 х 10 (А), 10 х 40 (Б-Д): скл — складчатость стенок, зо — зрелый ооцит, по — пристеночный ооцит, ро — растущий ооцит
Fig. 3. Sex tubules of the sea cucumber female from the Kiyevka Bay in June, 2011 (colored by hematoxylin-eosin): скл—plication of tubule walls, зo — mature oocyte, пo — wall oocytes, ро — growing oocyte. Magnification 10 х 10 (A), 10 х 40 (Б-Д)
нерации половых клеток (рис. 8). Встречались в отдельных трубочках сперматоциты I и II порядков. Аналогичная картина наблюдалась у трепанга из акватории у мыса Де-Ливрона (рис. 9).
Известно, что зимой у самок и самцов трепанга наряду со стадией накопления и дифференциации гамет, влекущей за собой увеличение размеров гонады, происходит зимняя резорбция половых клеток, которая, как правило, является необходимым условием для развития последующих генераций гамет (Низовская, 1971). Весной активизируются размножение и развитие половых клеток новой генерации.
Согласно нашим исследованиям, в бухтах Киевка, Мелководная и Соколовская нерест наблюдался с конца июля до конца первой декады августа. Массовый нерест трепанга в данном районе занимает более короткий промежуток времени — приблизительно одну декаду (первая декада августа). В третьей декаде сентября 2011 г. в бухте Киевка в полости тела трепанга находились мелкие трубочки с формирующейся

Рис. 4. Половые трубочки трепанга в июле 2011 г. в бухте Киевка (А-Г — вторая декада месяца, Д-Е — третья декада). Окраска гематоксилин-эозин. Ув. 10 х 40 (А-В, Д, Е), 10 х 10 (Г): А-В, Д, Е — самка, Г — самец; скл — складчатость, зо — зрелый ооцит, по — пристеночный ооцит, ро — резорбирующийся ооцит, сп — спермии
Fig. 4. Sex tubules of the sea cucumber from the Kiyevka Bay in middle July, 2011 (A-Г) and late July, 2011 (Д-E) (colored by hematoxylin-eosin): А-В, Д, Е — female, Г — male; скл — plication of tubule walls, зо — mature oocyte, по — wall oocytes, ро — resorption of oocyte, сп — sperms. Magnification 10 х 40 (А-В, Д, Е), 10 х 10 (Г)
складчатостью стенок, что может свидетельствовать о начале стадии восстановления (см. рис. 6). У трепанга из бухты Средней и у мыса Де-Ливрона в первой декаде октября в половых трубочках наряду со складчатостью уже шли процессы гаметогенеза: у самок имелись ооциты малого роста, а у самцов — сперматоциты I и II порядков (см. рис. 7-9). Учитывая вышеизложенное, можно предположить, что стадия покоя занимает один-полтора месяца (август — середина сентября).
Таким образом, в исследованный период (с мая по октябрь) у трепанга дальневосточного в половых железах протекали гаметогенетические процессы, характерные
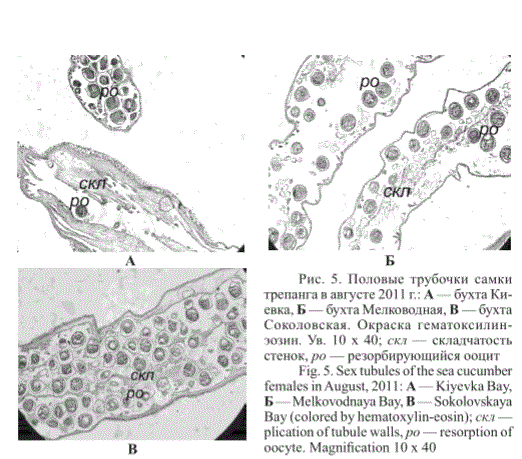
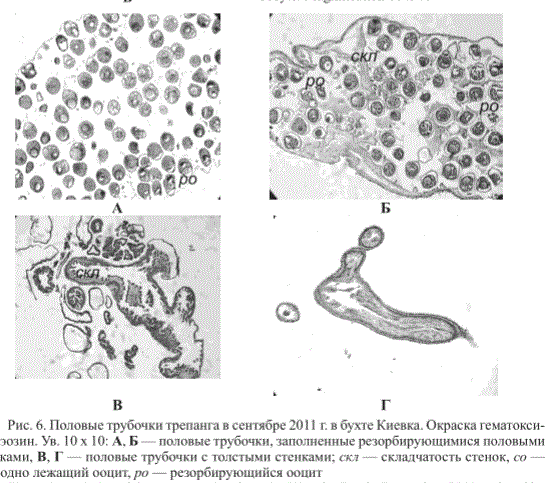
Рис. 6. Половые трубочки трепанга в сентябре 2011 г. в бухте Киевка. Окраска гематоксилин-эозин. Ув. 10 х 10: А, Б — половые трубочки, заполненные резорбирующимися половыми клетками, В, Г — половые трубочки с толстыми стенками; скл — складчатость стенок, со — свободно лежащий ооцит, ро — резорбирующийся ооцит
Fig. 6. Sex tubules of the sea cucumber from the Kiyevka Bay in September, 2011 (colored by hematoxylin-eosin): А, Б — sex tubules with resorption of oocyte, В, Г — sex tubules with thick walls; скл — plication of tubule walls, m — freely lying oocyte, ро — resorption of oocyte. Magnification 10 х 10
Рис. 5. Половые трубочки самки трепанга в августе 2011 г.: А — бухта Киевка, Б — бухта Мелководная, В — бухта Соколовская. Окраска гематоксилин-эозин. Ув. 10 х 40; скл — складчатость стенок, ро — резорбирующийся ооцит
Рис. 7. Половые трубочки самки трепанга в октябре 2011 г. в бухте Средней. Окраска гематоксилин-эозин. Ув. 10 х 10 (А), 10 х 40 (Б-Е): А — складчатость стенок трубочки, Б, Е — оогонии и растущие ооциты; ск — складчатость стенок трубочки, по — пристеночный ооцит, со — свободнолежащий ооцит
Fig. 7. Sexual tubules of the sea cucumber female from the Sredniaya Bay in October, 2011 (colored by hematoxylin-eosin): А — plication of tubule walls, Б, Е — oogonii and growing oocytes; ск — plication of tubule walls, по — wall oocytes, со — freely lying oocyte. Magnification 10 x 10 (A), 10 х 40 (Б-E)
для его репродуктивного цикла. Морфометрические параметры половых трубочек и соотношение гамет в них изменялись в зависимости от репродуктивного состояния особи.
С мая по октябрь 2011 г. у трепанга дальневосточного из бухт Киевка и Мелководная диаметр половых трубочек варьировал от 0,2 до 1,9 мм, диаметр свободнолежащих ооцитов изменялся от 71,4 до 167,0 мкм, диаметр пристеночных ооцитов — от 47,6 до 119,0 мкм. Гистологические картины у самцов и самок трепанга из бухт Мелководная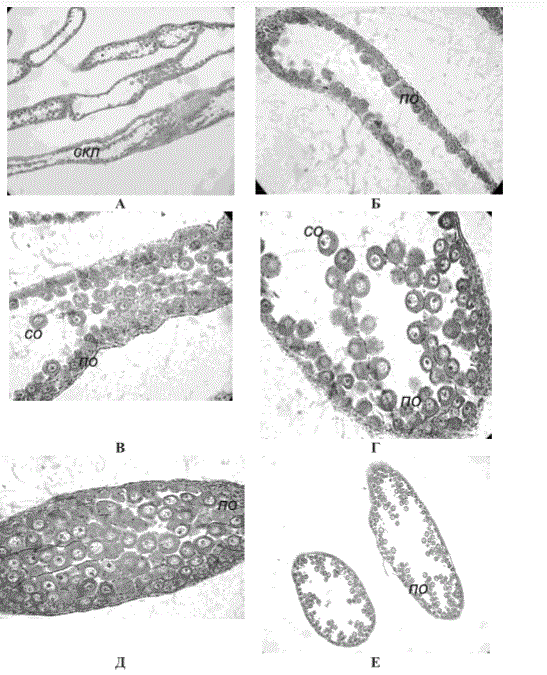
Рис. 8. Половые трубочки самца трепанга в октябре 2011 г. в бухте Средней. Окраска гематоксилин-эозин. Ув. 10 х 10 (А), 10 х 40 (Б): скл — складчатость стенок трубочки, сц — сперматоциты
Fig. 8. Sexual tubules of the sea cucumber male from the Sredniaya Bay in October, 2011 (colored by hematoxylin-eosin): скл — plication of tubule walls, сц — spermatocyte. Magnification 10 x 10 (A), 10 х 40 (Б)
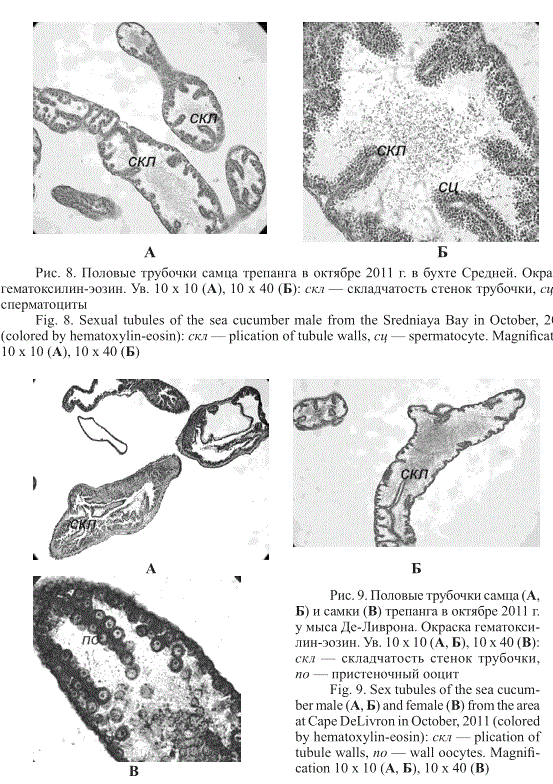
Рис. 9. Половые трубочки самца (А, Б) и самки (В) трепанга в октябре 2011 г. у мыса Де-Ливрона. Окраска гематоксилин-эозин. Ув. 10 х 10 (А, Б), 10 х 40 (В): скл — складчатость стенок трубочки, по — пристеночный ооцит
и Киевка в конце июля — начале августа свидетельствовали о готовности гонады к вымету гамет. Нерест у трепанга этих бухт наблюдался в первой декаде августа.
Наши исследования показали, что осенью наряду с резорбцией невыметанных зрелых клеток в половых трубочках активно идет процесс образования складчатости внутренней поверхности стенок и даже появляются новые генерации клеток, которые, скорее всего, в дальнейшем подвергнутся разрушению. В третьей декаде сентября 2011 г. в бухте Киевка в полости тела трепанга находились мелкие трубочки с формирующейся складчатостью стенок. В первой декаде октября в бухте Средней и у мыса Де-Ливрона в половых трубочках трепанга наряду со складчатостью уже
шли процессы гаметогенеза: у самок имелись ооциты малого роста, а у самцов — сперматоциты I и II порядков.
Стадия покоя, т.е. время, в течение которого не происходит формирования новых половых трубочек, занимает один-полтора месяца (август — середина сентября).
Выводы
Полученные данные по гистологической организации и клеточному составу половых желез трепанга дополняют сведения о его репродуктивной биологии и могут быть полезны для рационального ведения промысла этих животных и их воспроизводства.
Смотрите также






