Морфометрические различия
Эмбриональное развитие мальмы происходит в разнообразных условиях (Волобуев,
1975; Гриценко, 1990; Гудков, 1990; Черешнев и др., 2002). В верхнем течении на горных руслах
без перекатов в нересте участвуют жилая форма и карликовые самцы, в среднем и нижнем
течении — проходная и жилая формы, а также карликовые самцы. Генетическое единство
мальмы из бассейна р. Начилова подтверждает отсутствие достоверных различий по 9 мери-
стическим признакам между сеголетками из разных зон бассейна. При попарном сравнении
выборок из уловов станций Н3, Н4, Н7 и М3 только между сеголетками с разветвленных горных
(Н4) и равнинных (Н7) русел были получены достоверные различия по параметрам ll и vert.: в
среднем 138,7 (от 136 до 141) против 140,0 (от 138 до 142) чешуй в боковой линии и 65,5 (от 64
до 68) против 66,1 (от 65 до 68) позвонков соответственно. Величина различий минимальна
(0,05≤ р ≤0,01) и, по-видимому, связана с различным температурным режимом на нерестили-
щах (Павлов, 1989). Диапазон колебания величин всех признаков находится в пределах
изменчивости, характерной для вида (Черешнев и др., 2002).
Для сравнения двухлеток мальмы из разных зон по пропорциям тела использовали
выборки со станций Н3, Н4, Н6, Н7, Н8 и М3, выловленные в июле 2006 г. Небольшой объем
123
материала, собранного на горном русле без перекатов (Н3), не позволил исключить из анализа
мелкую молодь, т.е. при сравнении использовали одну выборку, особи которой отличались от
прочих достоверно меньшими размерами. Диапазоны изменчивости и средние значения
пластических признаков молоди из разных зон приведены в таблице 7.12.
Двухлетки мальмы ведут кочующий образ жизни (Савваитова, 1989; Гудков, 1990), от
чего величина различий по пластическим признакам между их выборками из разных зон
меньше, чем при сравнении выборок оседлого кижуча. Попарное сравнение индексов морфо-
метрических промеров, распределение которых было формально проверено на нормальность
(Джонгман и др., 1999), показало наличие достоверных различий между молодью по 5—12 из 25
сравниваемых пар (табл. 7.13). Объединение всех выборок в одну привело к потере симметрии
и одновершинности распределения 20 индексов. Нормальности распределения не утратили
лишь индексы, описывающие длину жаберной крышки (op), длину и ширину верхней челюсти
(lm и hmx), длину спинного (lD) и ширину анального (hA) плавников.
Наибольшие различия между выборками проявляются в диаметре глаза (о), высоте
головы и тела (hcz и H), длинах анального и грудных плавников (lA и lP), антивентральном
расстоянии (aV). Ширина лба (io), длина и ширина верхней челюсти (lm и hmx), длина спинного
(lD) и ширина анального (hA) плавников не различаются во всех парах сравнения.
При многофакторном разделении двухлеток мальмы, выловленных в разных зонах, в
функцию многомерной дискриминации включаются 13 из 25 индексов (асимметрия и эксцесс
распределений параметров незначимы). Дисперсия 5 из 13 дискриминирующих индексов дос-
товерно различается во всех группах сравнения. Наиболее значимыми разделяющими
параметрами являются признаки, которые проявляют максимальную изменчивость при попар-
ном сравнении: диаметр глаза (о), высота голова и тела (hcz и H), длина анального и грудных
плавников (lA и lP). Максимальное значение Лямбды Уилкса составляет 0,034. Объекты
(отдельные особи) включаются в состав групп, выделенных по зональному принципу, с
вероятностью 93 %, т.е уровень дискриминации выборок всего на 2 % ниже принятого уровня
достоверности.
Графическое отображение результатов сравнения методом канонического анализа ука-
зывает на относительно низкий уровень дискриминации выборок — факторные области
трансгрессируют (рис. 7.6 А). В наибольшей степени удалена от прочих область мальмы с
горных русел без перекатов, области выборок из уловов горных и равнинных русел с перека-
тами почти совпадают, эллипсоид устьевой мальмы перекрывает все прочие.
Необходимо отметить, что морфологическая обособленность двухлеток из верхнего
течения может быть связана не с формированием их внешнего облика в специфических
условиях на горных руслах без перекатов, а с аллометрическими особенностями роста, т.е. с
мелкими размерами особей в выборке. С целью выявления изменений пропорций тела
во время роста мальмы было проведено дополнительное сравнение выборок молоди разной
длины (табл. 7.14) из уловов участка р. Большая, примыкающего к устью р. Начилова (в самом
модельном водотоке необходимый объем выборок собрать не удалось).
Разноразмерную молодь из уловов участка р. Большая сравнивали по 16 индексам пла-
стических признаков, которые можно было измерить с высокой точностью у самых мелких
особей: c, hcz, lm, H, h, pl, lD, lA, lP, lV, aD, pD, aV, aA, P-V, V-A. Было установлено, что по кри-
терию Стьюдента выборки достоверно различались по 7 параметрам, причем только 2 из них
(lD и aD) различались во всех парах сравнения (0,01≤ р ≤0,001). Объединение 4-х выборок в
одну не нарушило нормальность распределения 12 индексов и только распределения индексов,
описывающих высоту головы (hcz), длину спинного плавника (lD), антидорзальное (aD) и вен-
тро-анальное (V-A) расстояния приобрели многовершинный вид. Следовательно,
аллометрические различия у обследованной молоди мальмы меньше, чем при сравнении
выборок из разных зон бассейна р. Начилова, причем разноразмерная молодь в наибольшей
степени различалась по показателям, проявившим минимальную изменчивость при сравнении
молоди из разных зон (lD, aD и V-A). Таким образом, фенотипические различия между двухлет-
ками мальмы с горных русел без перекатов и двухлетками из среднего и нижнего течения
бассейна не могут быть объяснены аллометрическими различиями выборок.
124
Таблица 7.12. Морфометрическая характеристика мальмы 1+, выловленной в разных зонах бассейна р. Начилова в июле 2006 г. (индексы в % от
длины тела)
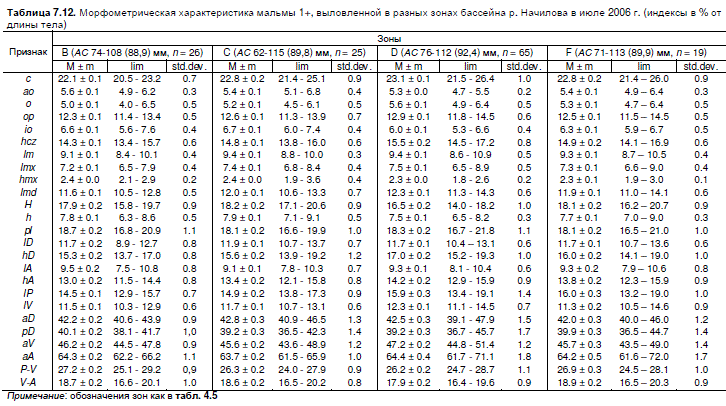
Таблица 7.13. Оценки достоверности различий между морфотипами мальмы 1+, выловленной
в июле 2006 г. в разных зонах бассейна р. Начилова, по критерию Стьюдента
Пары сравниваемых выборок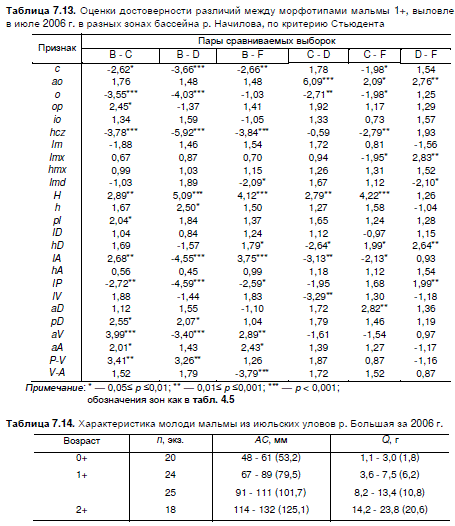 Примечание: * — 0,05≤ р
Примечание: * — 0,05≤ р
Сравнение морфометрических индексов одноразмерных двухлеток мальмы длиной 81—
111 мм, выловленных в пределах равнинного участка на станциях Н6, Н7 и М3 (выборки по 23—
25 экз.) в июле 2005 г., показало, что выборки из одной зоны достоверно различались по 6 из 25
параметров, при этом лишь 1 индекс (о) достоверно различался во всех парах сравнения.
Многофакторное разделение выборок привело к включению в функцию дискриминации
6 параметров, дисперсия двух из них (о и pl) различалась во всех группах сравнения.
Максимальная Лямбда Уилкса составила 0,048. Объекты включались в состав выделенных
групп с вероятностью 64 %. Таким образом, фенотипическая неоднородность мальмы в
пределах зоны равнинных русел с перекатами значительно меньше, чем при сравнении молоди
из разных речных зон (рис. 7.6 Б).
126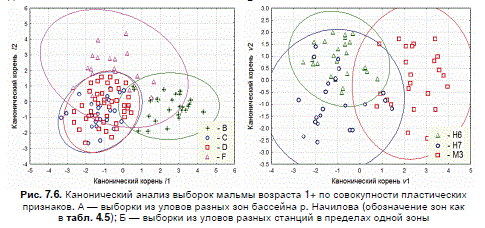
Рис. 7.6. Канонический анализ выборок мальмы возраста 1+ по совокупности пластических
признаков. А — выборки из уловов разных зон бассейна р. Начилова (обозначение зон как
в табл. 4.5); Б — выборки из уловов разных станций в пределах одной зоны
равнинных русел с перекатами.
В соответствии со сменой зон в бассейне р. Начилова у пестряток массовых видов лосо-
севых рыб с продолжительным пресноводным периодом жизни происходит смена
пространственных группировок, различающихся чертами внешней морфологии. В июле от исто-
ков до устья реки друг друга сменяют 3 группировки двухлеток кижуча и 2 группировки мальмы.
В устьевой зоне, судя по размаху изменчивости всех признаков, группировка включает как
местную молодь со специфическими признаками, так иммигрантов, скатившиеся из
вышележащих зон и поднявшиеся из главной реки.
Результаты изучения морфо-биологических особенностей молоди дают основание
считать, что внешняя морфология рыб помимо наследственных факторов в значительной
степени формируется под влиянием специфических условий среды. На фоне отсутствия досто-
верных различий по меристическим признакам двухлетки кижуча и мальмы из разных зон
достоверно различаются по 5—19 из 25 пластических признаков, а также по скорости роста,
микроструктуре и темпам нарастания регистрирующих структур. Во внешней морфологии наи-
большие различия получены по признакам, характеризующим положение и длину плавников,
высоту тела, диаметр глаза. По-видимому, это связано с особенностями локомоции и ориента-
ции в потоке, характер которого различается в разных зонах. Так, если на горных руслах с
мощным потоком эффективнее иметь низкое тело и короткие парные далеко расставленные
плавники, то на равнинных руслах — высокое тело и длинные парные плавники. Совершенно
очевидно, что высокая морфо-экологическая пластичность молоди на организменном уровне
позволяет лососевым рыбам эффективно приспосабливаться к внешним условиям и более
полно использовать ресурсы среды на всем протяжении речного бассейна. Полученные мор-
фологические различия между двухлетками кижуча и мальмы из разных зон выше
внутризональных и межгодовых различий, а также различий между молодью разного размера с
одного участка, что также подтверждает справедливость выделенных пространственных груп-
пировок.
Методически важно, что в связи с обнаруженными фенотипическими различиями между
молодью лососевых рыб внутри одной популяции, описания пластических признаков видов в
биотопически разнородной среде следует давать с учетом пространственной изменчивости
этих признаков. В противном случае можно получить достоверные различия между выборками
молоди соседних рек, определяемые спецификой условий обитания на руслах разных типов, а
не биологическими особенностями популяций этих рек.
Смотрите также






