Кета Oncorhynchus keta (Walbaum)
Популяционная структура
Этот вид лососевых рыб представлен множеством локальных популяций, различающихся рядом биологических особенностей, в том числе экологией размножения. Локальные популяции кеты репродуктивно изолированы, что установлено на основании анализа ряда генетических признаков и, в первую очередь, частот аллелей локусов некоторых ферментных систем [Бирман, 1964, 1977; Воловик, Ландышевская, 1968; Иванков, 1970, 1972; Куликова, 1970; Алтухов, 1974, 1983; Клоков, 1975; Николаева, 1978; Волобуев, 1983а, 1986; Okazaki, 1982a,b; Beacham et al., 1985]. Морфометрические и остеологические различия между популяциями кеты незначительны и не дают оснований для таксономического разделения [Горшков, 1979; Черешнев, 1980].
Основоположником изучения популяционнои структуры кеты является Л.С. Берг [1953], описавший в бассейне Амура летнюю и осеннюю расы, различающиеся сроками нерестового хода, сроками и местами нереста, размерами тела и плодовитостью. Дальнейшие исследования показали, что между летней и осенней кетой существует высокоэффективная репродуктивная изоляция. Развитие эмбрионов летней кеты происходит в подрусловом потоке, к которому примешаны грунтовые воды, развитие эмбрионов осенней кеты — в грунтовых водах [Леванидов, 1968]. Установлено, что летняя и осенняя расы кеты различаются числом хромосомных плеч [Куликова, 1971].
С момента описания Л.С. Бергом сезонных рас кеты, размножающейся в бассейне Амура, многие исследователи, изучающие кету, пытались отождествить выделение ими популяции с той или иной сезонной расой. А.И. Смирнов [1975] считает такой подход неверным. По его мнению, экологическое и морфологическое разнообразие кеты определяется не только различным сезоном размножения, но и привязанностью в период эмбриогенеза как к водам подруслового потока, так и к грунтовым водам. Эти воды значительно различаются температурным режимом, газовым и солевым составом. При этом в разных регионах — крупных речных системах, изолированных с древнейших времен, в один и тот же сезон размножаются экологически различные формы. Они, в свою очередь, разделяются на локальные стада или популяции, которые могут быть также дифференцированными. Подобный подход нам представляется обоснованным. Аналогичное мнение высказывают P.M. Викторовский и соавторы [1986]: "сезонные формы развились, по-видимому, на разной генетической основе, имеют различные уровни обособленности, и попытки рассматривать их как особый универсальный уровень популяционнои структуры не имеют под собой твердого основания".
По имеющимся в настоящее время данным, едва ли возможно представить родственные отношения между популяциями кеты и место каждой из них в популяционнои иерархии вида. Отсутствие знаний достаточно большого количества ситуаций не позволяет представить картину в целом. Так, в работе Е.А. Салменковой и соавторов [1986], подводящей итоги многолетних исследований генетической структуры кеты тихоокеанского побережья России, количественная оценка генетической обособленности популяций отдельных районов (например, Сахалино-Курильского) дает убедительную картину, согласующуюся с представлениями, полученными негенетическими методами. В то же время, по исследованным авторами признакам генетически близкими оказываются географически весьма удаленные и репродуктивно заведомо обособленные популяции (например, рек восточной Камчатки и материкового побережья Охотского моря). В этой связи следует с известной осторожностью интерпретировать данные о большой генетической близости амурской летней и осенней кеты, приведенные в упомянутой работе, поскольку они находятся в значительном противоречии с тем, что известно о биологии этих форм.
В качестве рабочей гипотезы мы принимаем, что вид Oncorhynchus keta разделен на ряд экологических форм, различающихся сроками размножения и использованием для нереста грунтовых вод и вод подруслового потока. Ареалы этих форм широко перекрываются. Форма, доминирующая в одном из регионов, может быть представлена в виде множества небольших популяций, мозаично распространенных в ареале другой формы. Предварительно можно считать, что таких форм существует четыре: летняя кета, откладывающая икру в воды подруслового потока (амурская летняя кета) [Берг, 1948; Леванидов, 19681; летняя кета, откладывающая икру на выходах грунтовых вод (камчатская летняя кета [Смирнов, 1975]; осенняя кета, откладывающая икру в воду подруслового потока (камчатская осенняя кета "манок" [Абрамов, 1948]; осенняя кета, откладывающая икру на выходах грунтовых вод (амурская осенняя кета) [Берг, 1948; Леванидов, 1969]. Каждая из этих форм представлена множеством иерархически подразделенных популяций. Число уровней в той или иной популяции, видимо, не одинаково и зависит от размеров и гидрографической подразделенности населяемых водоемов.
Распространение осенней кеты, нерестящейся в местах выхода грунтовых вод, как показали недавние исследования, гораздо шире, чем считалось раньше. Она в небольших количествах обнаружена в реках материкового побережья Охотского моря [Волобуев, 1983а, 1986]. Осенняя кета, эмбрионы которой развиваются в подрусловом потоке, известна только на юге Камчатки.
На Сахалине кета представлена летней и осенней формами, аналогичными амурской летней и осенней кете. Это представляется нам вполне закономерным, так как реки Сахалина являются остатками системы Палеоамура [Линдберг, 1972]. Осенняя кета распространена по всему охотоморскому побережью острова. На северо-западном побережье она встречается во всех реках от северной оконечности острова до пролива Невельского. Наиболее крупные стада воспроизводятся в реках Большие Лангры, Чингай и Большая. По восточному побережью осенняя кета входит практически во все реки от мыса Елизаветы до мыса Терпения, в заливе Терпения — в Поронай, в южных районах Сахалина популяции кеты существуют в реках Найба и Лютога. Искусственно воспроизводимая популяция осенней кеты есть в реке Ударница (бассейн озера Тунайча). В районе побережья Японского моря осеннюю кету искусственно воспроизводят в бассейнах рек Калининка, Заветинка и Ясноморка. В другие реки этого района осенняя кета заходит единично, и эти заходы едва ли можно рассматривать как свидетельство существования природных популяций. Наиболее крупными являются стада осенней кеты рек Тымь, Поронай, Найба, Набиль, Большая Лангры.
В.Н. Иванков [1972] по срокам хода, плодовитости, размерному и возрастному составу нерестовых стад, а также по степени выраженности брачного наряда при заходе в реки выделяет четыре группы популяций осенней кеты Сахалина: популяции бассейна Тыми и, видимо, более мелких рек северо-восточного побережья; популяции бассейна Пороная и прилежащих рек; популяции юго-восточного побережья; популяции югозападного побережья. Сходные результаты были получены А.А Ковтун [1983] на основании анализа структуры чешуи. По данным Ф.Нива и соавторов [Neave et al., 1976], кета северного и южного Сахалина различается районами морского нагула. Между стадами сахалинской кеты, воспроизводящейся в различных реках, обнаружены значительные генетические различия [Алтухов, 1974; Алтухов и др., 1983; Салменкова и др., 1986].
Анализ популяционной структуры кеты Сахалина осложняется многочисленными перевозками икры из одних рек в другие при проведении рыбоводных работ. Наименее подвержены влиянию перевозок стада рек Тымь и Калининка. Картина перевозок получается довольно пестрой, однако в ней можно выделить два основных направления: из Тыми в Поронай и в небольшом количестве в реки южного Сахалина; из Калининки в реки южного Сахалина [Рухлов, 1983].
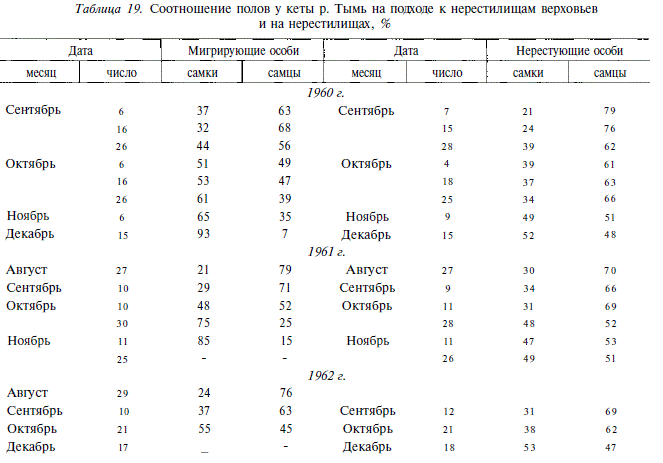
У осенней кеты в небольших сахалинских реках, таких, например, как р. Калининка, на основании различий в частотах аллелей локусов ряда ферментных систем у производителей разных сроков хода выделялись "простейшие популяционные единицы" [Алтухов, 1974]. В этой связи интересно рассмотреть, как изменяется у кеты соотношение полов среди мигрантов и нерестующих особей.
Известно, что у тихоокеанских лососей соотношение полов не остается постоянным на протяжении нерестового хода. Как правило, в начале хода преобладают самцы, в конце — самки, в середине хода соотношение полов приблизительно 1:1 [Агапов, 1941; Foerster, Ricker, 1953 и др.]. Эта особенность лососей используется при прогнозировании интенсивности и продолжительности их подходов в зону действия ставных неводов [Ефанов и др., 1978].
Иным бывает соотношение полов на нерестилищах, там на протяжении нереста преобладают самцы, т.е. самцы дольше самок задерживаются на нерестилищах и способны неоднократно нереститься со вновь подходящими самками [Кузнецов, 1937; Чебанов, 1983]. В 1960-1962 гг. мы регулярно на протяжении нерестового хода определяли соотношение полов у мигрирующей и нерестующей кеты. При этом старались, чтобы разрыв между сроками определения соотношения полов у мигрантов и нерестящихся особей не превышал 2 сут. От начала к концу хода среди мигрантов закономерно уменьшается доля самцов (табл. 19). В конце ноября — декабре подходы на 80-90 % представлены самками. На нерестилищах же самцы доминируют на протяжении всего нереста, и лишь в конце его соотношение полов выравнивается. Наибольшим преобладание самцов на нерестилищах бывает в начале нереста (в конце августа — первой половине сентября) и в конце массового нереста (во второй половине октября). Большая длительность нереста у самцов, чем у самок, зашедших на нерестилища одновременно с ними, способность неоднократно нереститься со вновь подошедшими самками приводят к смешиванию производителей разных сроков хода. Это явление должно препятствовать возникновению внутри хода репродуктивных групп, изолированных во времени, что дает основание рассматривать всех производителей, идущих в тот или иной нерестовый приток или небольшую реку, в качестве препродуктивного единства. В связи с этим вызывает сомнение правомерность выделения нескольких простейших популяционных единиц кеты в небольших сахалинских реках на основании различий генетических признаков у производителей разных сроков хода. Тем более что, как уже говорилось выше, на примере нерки показано, что изменчивость по локусам лактатдегидрогеназы и фосфоглюкомутазы имеет селективную природу. При этом направление отбора меняется на протяжении жизни генерации. Какова бы ни была физическая природа факторов отбора, воздействие их не может быть одинаковым из года в год, не может быть одинаковым на все группы особей простейших популяционных единиц. Это обстоятельство наряду с различиями в сроках ската молоди, широким пространственным распределением в море в период нагула, разной продолжительностью жизни особей, изменением соотношения полов и возрастных групп в процессе нерестового хода, различным соотношением возрастных групп в пределах полов может обусловить довольно пеструю картину изменчивости частот аллелей на протяжении нерестового хода производителей, которая не обязательно должна быть связана с популяционной подразделенностью.