Особенности морфологии и поведения камчатского краба
Начатые в 1995 г. в Баренцевом море специализированные планктонные съемки, направленные на сбор личинок камчатского краба, позволили получить первые данные по срокам выклева личинок, их распределению и численности (Баканев, Кузьмин, 1999; Беренбойм, 2001; Матюшкин, Ушакова, 2002). Однако до настоящего момента изучение морфологии ранних стадий развития акклиматизированного в Баренцевом море камчатского краба не проводилось. Детально изучить развитие крабана всех этапах раннего онтогенеза стало возможным только при выращивании в аквариальных условиях.
Презоэа (рис. 4.1 А) в основном сосредотачиваются на днебассейна. Основные морфологические особенности, обнаруженные нами у презоэа (Борисов, Ковачева, 2003; Ереlbaum et al., 2006) — наличие эмбриональной кутикулы, покрывающей как чехлом все тело личинки и образующей мягкие полые выросты на антеннах и тельсоне, — описаны и для презоэа других видов декапод [Konishi et al. 1987; Hong, 1988;; Guerrao, Abello, 1996]. Ha aHTeHHax I, II M. TeJIEcoHe KyTMкула образует мягкие полые перистые выросты с тонкими стенками (рис. 4.1 Б, E). Конечные щетинки презоэа имеют такое же строение, как конечные щетинки зоэa I, однако они покрыты эмбриональной кутикулой и большинство щетинок втянуты (рис. 4.1 Г, Д). На стадии презоэа отсутствуют рострум и постеролатеральные шипы на 4 сомите абдомена, на карапаксе имеются дорзо-латеральные шипы.
Личинки выклевываются с ввернутыми щетинками, а наличие выростов эмбриональной кутикулы позволяет презоэа перемещаться в толще воды. Таким образом, личинки имеют возможность рассредоточиться сразу после выклева.
Другое объяснение в поддержку интерпретации презоэа крабов Porcellanidа как естественной стадии, а не лабораторного артефакта (как предполагали ранее) предлагалось С.Л. Гонор и Ж. Ж. Гонор (Conor, Gonor, 1973): коротко живущая, округлая личинка с немногими выростами тела способна более эффективно избегать запутывания частичек детрита между щетинками и выростами сразу после выклева.
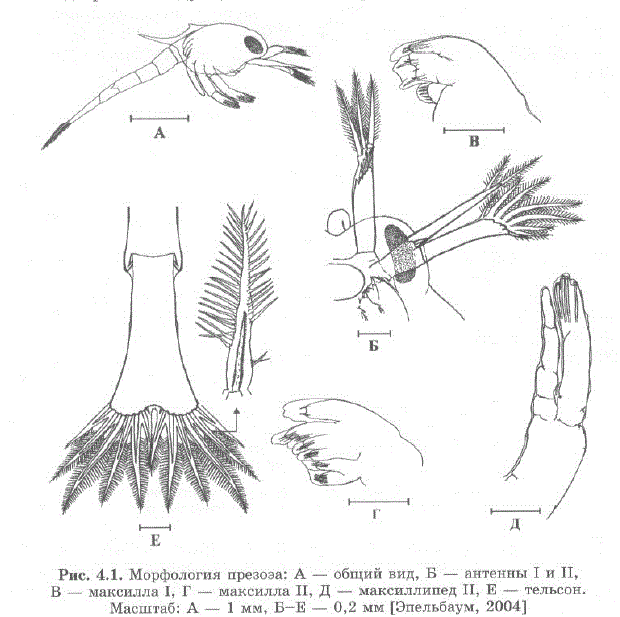
Рис. 4.1. Морфология презоэa: A — общий вид, Б — антенны I и II, В — максилла I, Г — максилла II, Д — максиллипед II, E — тельсон. Масштаб: A — 1 мм, Б—E — 0,2 мм (Эпельбаум, 2004)
По нашим наблюдениям, презоэа периодически поднимаются в толщу воды и снова медленно опускаются на дно (Борисов, Ковачева, 2003; Ковачева и др., 2005а). Для движения презоэа использует перистые кутикулярные выросты и плавает, совершая хлопающие движения абдоменом (быстро сгибая и разгибая его), при этом тельсон играет роль гребной лопасти. Кутикулярные выросты антенн I и II, раскрываясь наподобие веера, замедляют погружение личинки, когдата прекращает движения абдоменом (Эпельбаум, 2004).
Эксперименты, проведенные нами при температуре воды 7—8 С., показали, что стадия презоэа чаще всего длится менее часа. Линька происходит быстро, в течение нескольких минут эмбриональная кутикула сбрасывается, выворачиваются щетинки на конечностях личинки (Kovatcheva et al., 2006b). У личинок, не перелинявших в течение полутора часов с момента вылупления, отмечались многочисленные уродства: недоразвитые щетинки на конечностях, деформированный рострум и т.д. И, хотя такие личинки существовали еще более 10 дней, они не могли нормально плавать и питаться, были малоактивны и, в конечном итоге, погибли, имея разную степень морфологических отклонений (рис. 4.2).
Рис. 4.2. Морфологические отклонения презоэа: A — MaKcMJIJIa I, B — MaKcMJIJIa II. LIIKaJIa: 0,2 MM
Недоразвитие щетинок на конечностях пищедобывательного аппарата (см. рис. 4.1 В, Г) и наличие эмбриональной кутикулы свидетельствуют о том, что презоэа не питается (Эпельбаум, 2004; Ковачева и др., 2005a].
После линьки презоэа превращается в зоэа, которая также как и презоэа морфологически сильно отличается от взрослого краба (рис. 4.3).
Рис. 43. Зоза |
Морфологические различия личинок крабов подотряда Anomura описаны С. Сато (Sato, 1958) и Р.Р. Макаровым (1966). На основании этих данных В.С. Левин [2001] выделил отличительные признаки I—IV стадий зоэа камчатского краба. В данной работе мы приводим более полную и несколько модифицированную нами схему для определеHMSI cTaII Mii. 303a Paralithodes camtschaticus (Tao.J.I. 4.1). B IIpe IIJIO2KeHной нами схеме учитывается не только наличие плеопод, уропод и обособленность тельсона (рис. 4.4), но также и щетиночное вооружение экзоподитов максиллипед третьей пары — критерий, позволяющий легко отличить зоэa I от зоэa II (рис. 4.5) (по схеме В.С. Левина разделение этих стадий затруднительно) (Kovatcheva, Epelbaum, 2003; Ковачева и др., 2005в.
Таблица 4.1. Отличительные признаки зоэа камчатского краба I—IV стадий развития
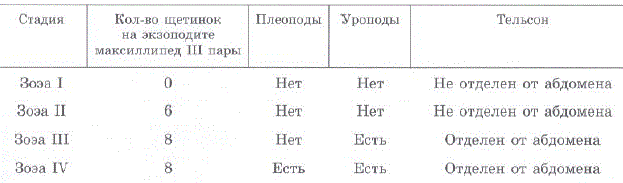
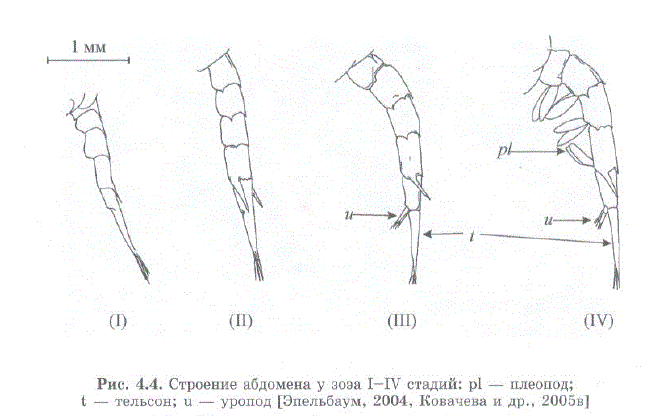
Рис. 4.4. Строение абдомена у зоэa I—IV стадий: pl — плеопод; t — тельсон; u — уропод (Эпельбаум, 2004, Ковачева и др., 2005 в)
Участок гребной щетинки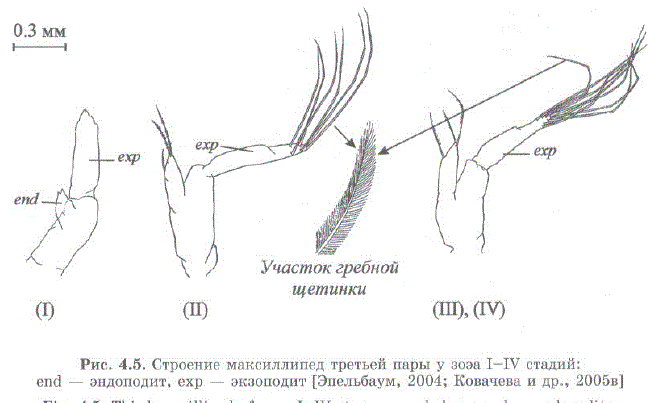
Рис. 4.5. Строение максиллипед третьей пары у зоэa I—IV стадий: end — эндоподит, exp — экзоподит (Эпельбаум, 2004; Ковачева и др., 2005 в)
Общая морфология частей тела и придатков, по существу, остается неизменной в течение всего личиночного периода. С каждой линькой личиночные придатки становятся пропорционально больше, и некоторые из них приобретают дополнительные щетинки и/или сегменты и части. Расположение придатков, которые могут быть вовлечены в процесс добычи пищи, показано на рис. 4.6.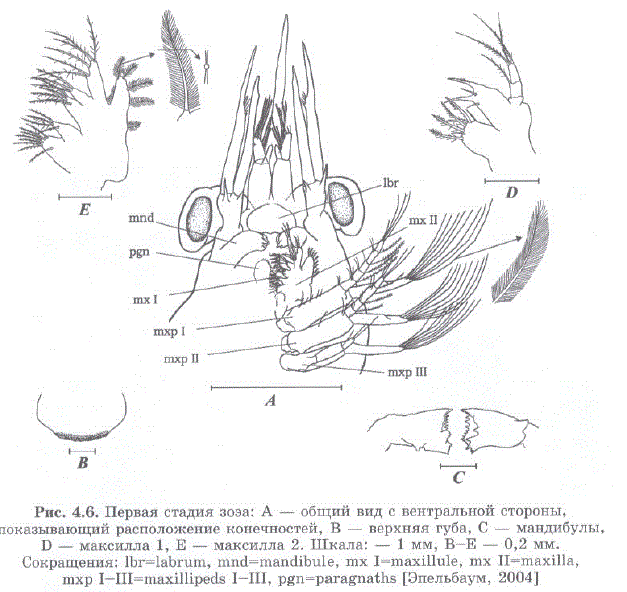
Рис. 4.6. Первая стадия зоэa: A — общий вид с вентральной стороны, показывающий расположение конечностей, B — верхняя губа, C — мандибулы, D — MaKcMJIJIa 1, E — MaKCMJIJIa 2. IIIKaJIa: — 1 MM, B—E — 0,2 MM. CokpaIILeHMSI: libr=labrum, mind-mandibule, mix I=maxillule, mix II=maxilla, mXp I-III=maxillipeds I-III, pgn=paragnaths 9IIe.JIL6ay M, 2004
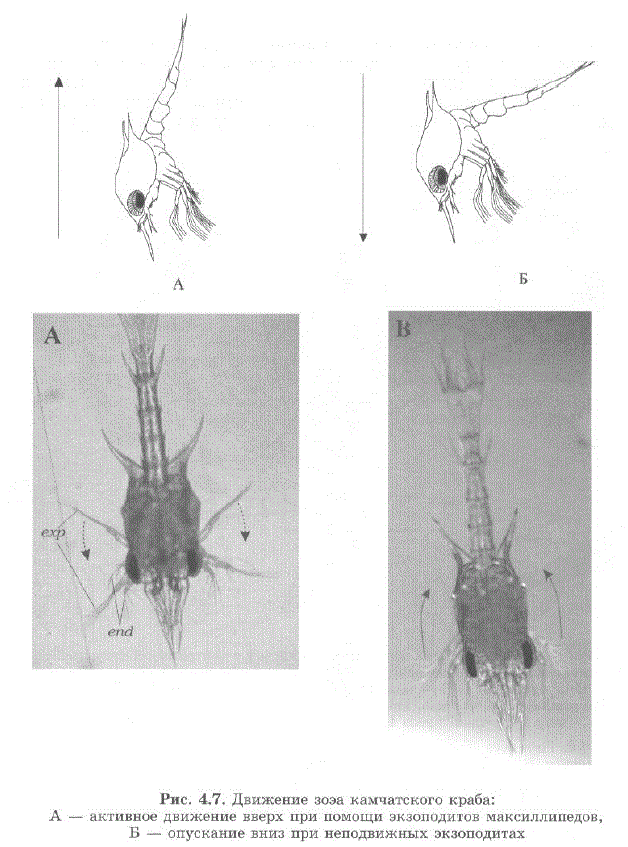
Личинки I—IV стадий ведут планктонный образ жизни. После перехода на стадию зоэa I, способ движения личинки изменяется. Локомоторную и вододвигательную функцию узоэа выполняют экзоподиты максиллипед, перистые щетинки на концах которых образуют гребные лопасти, вогнутые с оральной стороны (см. рис. 4.6 A).
Гребной удар создает течение, направленное вперед к рострумуличинки. Работая экзоподитами, личинка плывет тельсоном вперед, поднимаясь вверх к поверхности воды, занимая при этом вертикальHoe IIOJIO 3 KeHMe pOCTpyMOM BHIM3; IIp M. HeIIOJI BIM2KIHB IX 9K3OI IOJIMTaX JIMчинка опускается вниз. Личинка может координировать направление движения, изменяя положение максиллипедительсона. Эндоподиты максиллипед остаются в вытянутом состоянии и не принимают участия в плавании (рис. 4. 7 А, Б).
Выявленные нами функциональные особенности морфологии личинок Р. сатschatiсиs были использованы при разработке, в частности, способов кормления, размера кормовых частиц, конструктивных особенностей установок для культивирования, учитывающих определяемое морфологическим строением поведение (Эпельбаум, 2004; Ковачева, 2005, Ковачева и др., 2005а, в Ереlbaum, Kovatcheva, 2005, Kovatcheva et al., 2006; Epelbaum et al., 2006b).
Личинки и глаукотоэ выживают при солености 27—40%о [Nakanishi, 1987; Орлов, 1998]. При воспроизводстве камчатского краба в искусственных условиях соленость поддерживали в пределах 32—35%o, что соответствует диапазону колебаний солености в природе, в районах обитания краба у побережья Тихого океана и Баренцева моря.
Смотрите также
