Послеличиночной период - Особенности морфологии и поведения
После линьки зоэa IV превращается в послеличинку — глаукотоэ. Внешне глаукотоэ уже похожа на взрослого краба, имеет развитые перейоподы (рис. 5.1 Б, В) и покрытый шипами карапакс, однако абдомен остается длинным, как уличинки (рис. 5.1 A). Плавательный аппарат представлен хорошо развитыми плеоподами (рис. 5.1 Г), поэтому, в отличие от зоэа, глаукотоэ плавает рострумом вперед. Первые 3—4 суток в течение дня глаукотоэплавают в толще воды, а к вечеру опускаются на дно и удерживаются клешненосными перейоподами на частичках субстрата.
Такое поведение связано с тем, что в природе основной функцией глаукотоэ является поиск субстрата для оседания, который обеспечит будущих мальков питанием и защитой. Некоторые глаукотоэ практиLLLLaaaLLLLL LLL LLGLGLL0LLLLLLLS LLLLLLaaGGL LLaaLLLLLLGLLLLLLLLGLLLGLLLLL0L L LL LLLGLLLLLLL 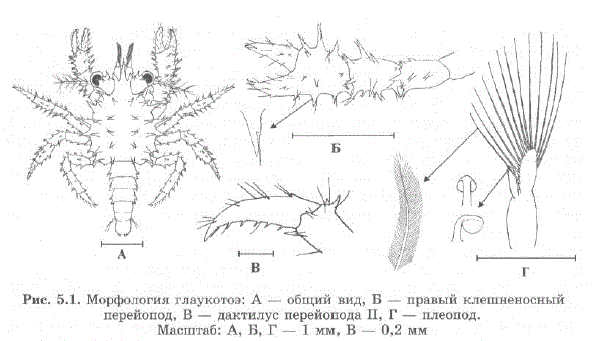
Рис. 5.1. Морфология глаукотоэ: A — общий вид, Б — правый клешненосный перейoпод, В. - дактилус перейoпода II, Г - плеопод. MacIIITa6: A, B, T – 1 MM, B – 0,2 MM
ночного периода Ковачева и др., 2005 в Коvatcheva et al., 2006b, Epelbaum et al., 2007).
У глаукотоэмаксиллипеды относительно малы, а их экзоподиты имеют слаборазвитые гребные лопасти. По нашим наблюдениям, в аквариальных условиях глаукотоэ не питаются. При содержании без пищи они благополучно прошли через процесс линьки и превратились в мальков (Ковачева, 2002а; Эпельбаум, 2002; Ковачева и др., 2005B; Epelbaum et al., 2006)
Морфологический анализ пищедобывательного аппарата подтвердил, что мандибулы и максиллипеды не приспособлены для обработки пищи. Так, конечности глаукотоэ малы, их щетиночное вооружение недостаточно развито и малопригодно для обработки пищи (рис. 5.2).
Анализ строения пищеварительной системы глаукотоэ и ее сравнение с пищеварительной системой нормально питающейся зоэа также указывают на то, что она не питается. В желудке глаукотоэслабо развит фильтрационный пресс, на стенках желудка отсутствуют какиелибо щетинки или зубчики, выросты гепатопанкреаса и дивертикулы средней кишки значительно меньше по размеру (рис. 5.3). В клетках эпителия пищеварительной железы и дивертикулов средней кишки личинки на стадии зоэа имеются липидные капли [Abrunhosa, Kittaka, 1997a; Эпельбаум, 2002; Эпельбаум, 2004; Ковачева и др., 2005в). Можно предположить, что накопленные на стадии зоэа, они обеспечивают непитающуюся глаукотоэ энергетическими ресурсами.
Средняя кишка и главная доля гепатопанкреаса глаукотоэ относительно длиннее, чем на стадии зоэа (рис. 5.3). Это обеспечивает увеличение площади контакта между гепатопанкреасом и гемолимфой, приводящее к более эффективной передаче энергетических ресурсов [Abrunhosa, Kittaka, 1997а; Эпельбаум, 2004; Ковачева и др., 2005B; Epelbaum et al., 2006.
По нашим наблюдениям, для глаукотоэ характерно наличие тонких сенсилл на всех шипах карапакса и перейопод (см. рис. 5.1 Б) и даже на шипиках глаз. Подобные волоски, связанные с сенсорными окончаниями, имеются у многих артропод и являются единственным местом контакта эпителиальных слоев с внешней средой (Shelton, Laverack, 1970). По-видимому, эти сенсиллы являются механо- и хеморецепторами и у глаукотоэ играют важную роль при выборе субстрата для оседания.
Каковы же возможные причины афагии глаукотоэ? Основная функция глаукотоэ — поиск микробиотопа, который сможет обеспечить будущих мальков защитой и питанием (Freese, Babcock, 1989; Stevens, Kittaka, 1998]. Mo}KHO IIpe IIIIOJIO2KMTE, HTO OTCyTCTBMe Heo?xoдимости поиска пищи позволяет глаукотоэ совершать более длительные локальные миграции для поиска подходящих условий. Глаукотоэ может продолжать вести планктонный образ жизни или немедленно осесть при обнаружении удобного микробиотопа (Strathman,
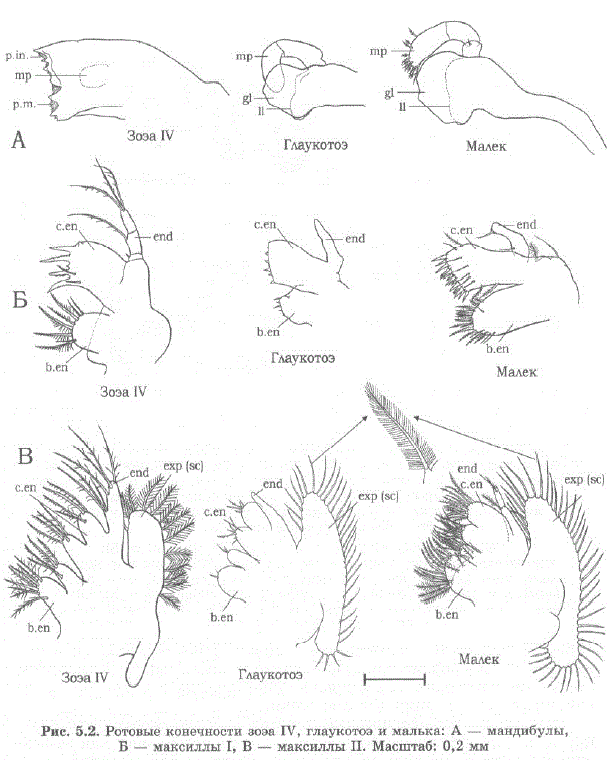
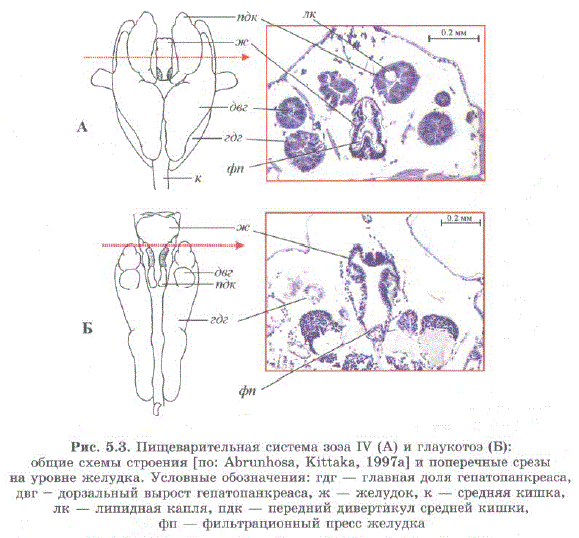
Рис. 5.3. Пищеварительная система зоэa IV (А) и глаукотоэ (Б): общие схемы строения [по: Abrunhosa, Kittaka, 1997a] и поперечные срезы на уровне желудка. Условные обозначения: гдг — главная доля гепатопанкреаса, двг — дорзальный вырост гепатопанкреаса, ж — желудок, к — средняя кишка, лк — липидная капля, пдк — передний дивертикул средней кишки, фп — фильтрационный пресс желудка Fig. 5.3. Digestive system of zoea IV (A) and glaucothoe (B): general scheme of structure Abrunhosa, Kittaka, 1997a) and cross section of stomach. Legend: TIK - main part of the hepatopancreas, IBT - dorsal hepatopancreatic incrustation, K - stomach, K - midgut, JIK - lipid drop, III JAK — front diverticulum of the midgut, q?II — filtrational press of stomach
1985]. Подобная мысль высказана P. Дейврс [Dawirs, 1981] в отношении непитающейся мегалопы рака-отшельника Pagитиs bernhardus: отсутствие необходимости в поиске пищи повышает шансы мегалопы найти пустую раковину для заселения.
Основные морфологические и поведенческие особенности глаукотоэ, которые мы рассматриваем как приспособление для эффективного расселения вида при поисках субстрата для оседания, отражены на рис. 5.4.
Наблюдения Б. Стивенс и Х. Китта на Stevens, Kittaka, 1998 в аквариумах показали, что глаукотоэ камчатского краба проявляют плавательную активность в дневные часы, будучи в значительной степени
Рис. 5.4. Особенности морфологии и поведения глаукотоэ Fig.
неподвижными между закатом и восходом солнца. В аквариумах со смешанными субстратами пропорция плавающих глаукотоэ в начале стадии была 28%, но уменьшалась до менее чем 10% к середине стадии, что хорошо согласуется с нашими данными о снижении двигательной активности камчатского краба к середине стадии глаукотоэ.
Смотрите также
