ИХТИОФАУНА МАЛЫХ ВОДОТОКОВ ЗАПАДНОЙ КАМЧАТКИ
Подавляющее число малых водотоков Западной Камчатки отличается низкой или уме-
ренно низкой температурой и мутностью воды, быстрым течением и каменистым ложем.
Подобные условия благоприятны для нереста, развития и нагула лососевых рыб из семейства
Salmonidae. К группе так называемых лососевых водотоков, где может проходить полный
цикл воспроизводства лососевых рыб (Леванидов, 1981; Леванидова и др., 1989), относятся
ручьи и малые реки региона — постоянные водотоки с собственной долиной, текущие по
разработанному руслу, продольный профиль которого выработан самим потоком, т.е. при усло-
вии, что мощности потока достаточно для перемещения и аккумуляция донных наносов (по
[Завадский, 2005]). Овражно-балочные водотоки на Западной Камчатке возникают в условиях
недостаточной мощности потока, их русла формируются за счет поверхностного размыва и
поступления внешних материалов (например, обвально-осыпных явлений, отложения
мелкофракционных материалов и т.п.). Переотложение наносов в руслах овражно-балочной
сети не выражено, аллювиальной толщи не формируется. Лососи, зарывающие свою икру в
грунт, способны использовать овражные водотоки лишь для нагула в летне-осенний период.
Рыбное население, т.е. совокупность рыб всех местных видов (Зданович, Криксунов,
2004) на Западной Камчатке отличается видовой обедненностью. Под этим понимается
несоответствие разнообразия таксонов рыб природным условиям региона, в котором, по дан-
ным ихтиогеографических сравнений, условия среды могли бы удовлетворить экологическим
запросам групп таксонов, населяющих другие районы со сходными природными условиями
(Голубцов, Малков, 2007). По современным оценкам на Западной Камчатке обитает не более
20—21 вида (Черешнев, 1996; Каталог позвоночных…, 2000; Федоров и др., 2003; Черешнев и
др. 2002), в ручьях и малых реках встречаются 16 из них (табл. 1.1). Оставшиеся виды при-
ходятся на обитателей солоноватых эстуариев и прибрежных озер: обыкновенная малоротая
корюшка Hypomesus olidus, морская малоротая корюшка Hypomesus japonicus, тихоокеанская
навага Eleginus gracilis, тихоокеанская сельдь Clupea pallasii и плоскоголовая широколобка
Megalocottus platycephalus.
Ихтиофауна подавляющего большинства малых рек полуострова характеризуется рез-
ким преобладанием эвригалинных и проходных рыб, причем 9 видов из последней группы
относятся к семейству Лососевые. Роль пресноводных видов минимальна, на Западной Кам-
чатке этот комплекс представлен всего двумя видами — истинно пресноводным камчатским
хариусом и реликтовой жилой формой арктического гольца Salvelinus sp. 4, который обитает в
оз. Начикинское (Черешнев и др. 2002; Савваитова, 1989).
В ихтиогеографическом отношении реки Западной Камчатки попадают в Южно-Камчат-
ский подокруг (на юг от бассейна р. Палана по охотскому побережью и р. Озерная-Восточная по
тихоокеанскому) в составе Охотско-Камчатского округа. Последний выделен в Тихоокеанской
провинции Циркумполярной подобласти в связи с историей формирования ихтиофауны (Берг,
1962; Черешнев, 1998). Считается, что рецентный комплекс видов, населявший реки полуост-
рова до последнего оледенения, закончившегося около 10 тыс. лет назад, был почти полностью
уничтожен (Линдберг, 1972; Леванидов, 1981). Основную часть современной фауны подокруга
сформировали рыбы из отряда Лососеобразные - Salmoniformes (лососи, форели, гольцы и
расселившийся из ледниковых рефугиумов хариус), представленные многочисленными внутри-
видовыми таксонами (Черешнев, 1998; Федоров и др., 2003).
Абсолютное доминирование лососеобразных рыб во многом объясняется их высокой
эволюционной пластичностью. Благодаря хомингу, т.е. возврату для размножения в родную
реку или даже на конкретный участок ее русла, популяции большинства видов из разных рек
оказываются в определенной степени изолированными, эволюционный процесс в них приобре-
тает собственную направленность (Коновалов, 1980; Алтухов и др., 1997). В сложных речных
системах (например, в крупнейшем западно-камчатском бассейне р. Большая) популяции могут
подразделяться на субпопуляции отдельных нерестовых притоков или озер (Бугаев, 1995;
Глубоковский, 1995).
7
Таблица 1.1. Фауна рыб ручьев и малых рек Западной Камчатки (латинские названия по
[Богуцкая, Насека, 2004])
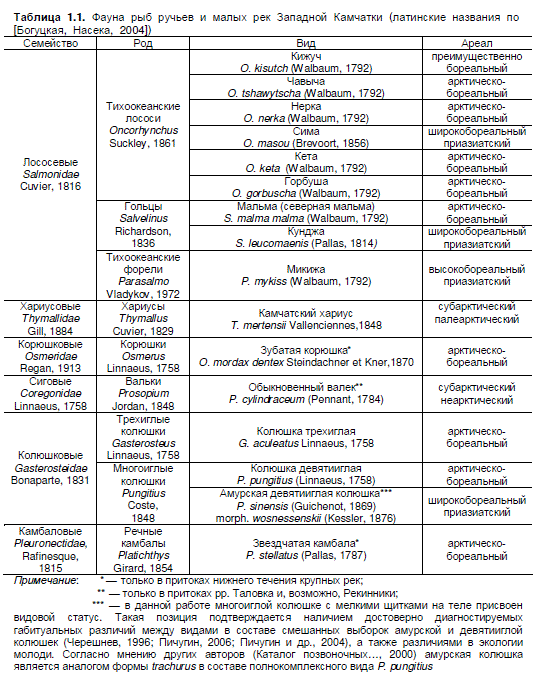
Примечание: * — только в притоках нижнего течения крупных рек;
** — только в притоках рр. Таловка и, возможно, Рекинники;
*** — в данной работе многоиглой колюшке с мелкими щитками на теле присвоен
видовой статус. Такая позиция подтверждается наличием достоверно диагностируемых
габитуальных различий между видами в составе смешанных выборок амурской и девятииглой
колюшек (Черешнев, 1996; Пичугин, 2006; Пичугин и др., 2004), а также различиями в экологии
молоди. Согласно мнению других авторов (Каталог позвоночных…, 2000) амурская колюшка
является аналогом формы trachurus в составе полнокомплексного вида P. pungitius
Структура субпопуляций усложнена образованием экологических форм (или морф), ко-
торые характеризуются различными жизненными стратегиями и могут существовать
автономно продолжительное время (Савваитова, 1989; Павлов, Савваитова, 2008). Разнообра-
зие жизненных стратегий подразумевает формирование нескольких вариантов
эпигенетического проявления линии развития (табл. 1.2). Формы различаются по комплексу
морфофизиологических признаков, возникающих в пределах нормы реакции в зависимости от
8
условий развития (Дгебуадзе, 1998). Внешняя среда действует как сигнал, включающий или вы-
ключающий соответствующий механизм, уже заложенный в геноме организма (Медников, 1987).
Таблица 1.2. Разнообразие типов жизненной стратегии лососеобразных рыб Западной
Камчатки
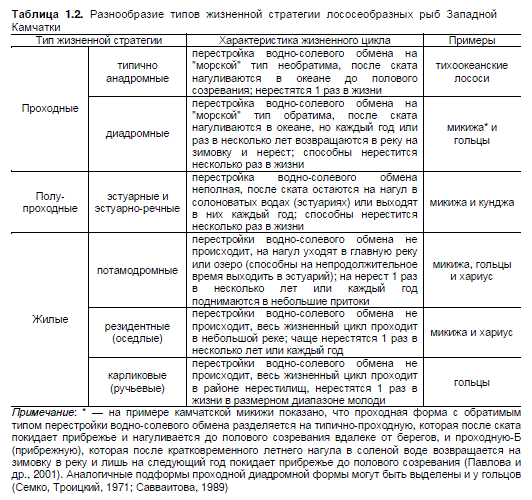
Примечание: * — на примере камчатской микижи показано, что проходная форма с обратимым
типом перестройки водно-солевого обмена разделяется на типично-проходную, которая после ската
покидает прибрежье и нагуливается до полового созревания вдалеке от берегов, и проходную-Б
(прибрежную), которая после кратковременного летнего нагула в соленой воде возвращается на
зимовку в реку и лишь на следующий год покидает прибрежье до полового созревания (Павлова и
др., 2001). Аналогичные подформы проходной диадромной формы могут быть выделены и у гольцов
(Семко, Троицкий, 1971; Савваитова, 1989)
Проходные лососи из одной популяции нередко разделяются на расы — группы особей,
имеющие стойкие морфологические особенности, передающиеся по наследству (Берг, 1962)
вследствие наличия специфических черт размножения, т.е. времени захода в реку, типа
избираемых нерестилищ, срока эмбрионального развития и т.п. (Никольский, 1953). Для
некоторых видов, например, для горбуши, образование рас связано с разделением популяции
на темпоральные группировки — группы особей, образованные потомками одного поколе-
ния (например, четных и нечетных лет). В отличие от форм, различия между расами в той
или иной степени генетически обусловлены (Атлантический лосось, 1998).
В ответ на высокую пространственно-временную неоднородность условий малых водо-
токов население молоди и взрослых жилых лососевых рыб разделяется на временные
функциональные группы — гильдии (Root, 1967) или агрегации в русскоязычной литературе
(Шилов, 1977; Живоглядов, 2004). Подобные элементы населения представляют собой неслу-
чайные группы особей на определенном этапе жизненного цикла, которые используют одни
и те же ресурсы в пределах участка водотока сходным образом (Wooton, 1998). Под ресур-
сами чаще всего понимают жизненное пространство и источники пищи (Glova, 1986; Li, Li, 1996).
Оседлые рыбы, которые на данном этапе жизненного цикла не покидают своего место-
обитания, т.е. входят в состав территориальных сообществ (Павлов, 1979), разделяются на
агрегации 4-х типов (по [Панов, 1983; Живоглядов, 2004]):
9
- агрегации пассивных резидентных особей — молодь в местах выхода из гнезд и рыбы всех
возрастов со сниженным уровнем метаболизма во время зимовки;
- вынужденные агрегации активных резидентных особей (пищевые скопления) — неустойчи-
вые скопления рыб в местах локального изобилия корма;
- территориальные агрегации активных резидентных особей — рыбы, поддерживающие
фиксированный пространственный контакт (удерживающие кормовой участок) в условиях лими-
тированной кормовой базы и высокой разнокачественности среды при относительно
стабильных внешних условиях (например, в летне-осенний период);
- агрегации нерестующих особей — половозрелые рыбы на нерестилищах;
Мигрирующие (в т.ч. участвующие в целенаправленном скате в соленые воды) и кочую-
щие рыбы, которые переселяются (перемещаются) с данного участка водотока вверх или вниз
по течению (Павлов, 1979; Павлов и др., 2007; Pavlov et al., 2005), разделяются на агрегации 4-х
типов (по [Панов, 1983; Павлов и др., 2006]):
- агрегации пассивно скатывающихся особей — покатники видов с коротким пресноводным
периодом жизни, ранняя молодь видов с продолжительным пресноводным периодом жизни во
время первичного расселения вниз по течению из районов нерестилищ, а также сносимая пото-
ком ослабленная молодь;
- активно-добровольные агрегации мигрирующий особей — рыбы на этапе вторичного рассе-
ления из притоков в главную реку или обратно;
- агрегации активно скатывающихся смолтов — сформированные смолты и рыбы, повторно
скатывающиеся после кратковременного нагула в соленой воде и последующей зимовки,
целенаправленно мигрирующие в соленые воды;
- активно-вынужденные агрегации мигрирующих особей — рыбы во время миграций из небла-
гоприятных местообитаний, где условия угрожают жизнедеятельности (например, при
промерзании участков реки), а также во время нерестовых миграций.
Количественное соотношение особей в составе временных агрегаций находится в тес-
ной зависимости от условий на данном участке водотока и определяет структуру его населения.
Так, сеголетки кижуча в августе в условиях достаточной обеспеченности пищей целиком вклю-
чаются в состав территориальных агрегаций резидентных особей, а при недостатке корма и
местообитаний разделяются на территориальных и пассивно мигрирующих (Chapman, 1962,
1966). Очевидно, что возможность изменения сочетания элементов в составе населения обес-
печивает пластичность способов эксплуатации окружающей среды (Шилов, 1977; Martel, 1996).
На Западной Камчатке в ручьях и малых реках нерестятся все виды лососевых рыб и
хариус. Вклад малых водотоков в воспроизводственный потенциал отдельных видов тихо-
океанских лососей был изучен на примере бассейна р. Большая, в котором водотоки короче
100 км занимают около 60 % водной поверхности (рис. 1.1). Из представленной таблицы, кото-
рая составлена на основе анализа фондовых материалов КамчатНИРО по аэровизуальным
учетам нерестилищ, выполненных Остроумовым А.Г. в 1950-х гг., видно, что в малых водотоках
сосредоточено около 40 % от бассейновых нерестилищ чавычи, 32 % нерестилищ кижуча,
21 % — реофильной нерки, 18 % — кеты и 15 % — горбуши (табл. 1.3). Основная часть нерес-
тилищ промысловых тихоокеанских лососей сосредоточена в 3-х главных реках длиной 100 и
более км (рр. Карымчина, Плотникова, Быстрая + Большая), около 5 % нерестилищ главных рек
приходится на небольшие лососевые (пойменные) ключи, являющиеся отшнуровавшимися ста-
ричными руслами. Распределение нерестилищ прочих лососевых рыб демонстрирует иную
картину. Так, все нерестилища микижи и ее проходной формы на Камчатке сосредоточены в
малых равнинных реках (Кузищин и др., 2002; 2006). В этих же водотоках нерестится бóльшая
часть кунджи (Черешнев и др., 2002) и камчатский хариус (Скопец, 1993). Сима поднимается на
нерест в предгорную зону малых рек и ручьев (Смирнов, 1975).
Важнейшую роль в воспроизводстве лососевых рыб малые водотоки играют за счет
использования их площадей молодью видов с длительным пресноводным периодом жизни для
нагула и зимовки. Известно, что молодь чавычи, кижуча, нерки, симы, микижи и гольцов во
время расселения с нерестилищ проникает в самые малые притоки, причем плотность обита-
ния в последних часто значительно выше, чем в главных руслах (Смирнов, 1975; Pacific
salmon…, 1995; Cote, 2007 и др.). Значение малых водотоков в воспроизводстве лососевых рыб
растет пропорционально увеличению густоты речной сети. На примере речных бассейнов
Северной Америки с густотой речной сети 0,55—0,65 км/км2 показана зависимость общей
продукции смолтов кижуча, чавычи и стальноголового лосося от количества малых рек (Bradford
et al., 2001; Sharma, Hilborn, 2001). Бассейн р. Большая отличается бóльшей разветвленностью
речной сети (0,67 км/км2) и, по аналогии, можно ожидать, что количество смолтов на выходе из
10
главной реки в большой степени определяется условиями обитания в малых реках и ручьях.
В конечном счете, обилие рыбного населения в бассейнах главных рек определяется
разнообразием условий обитания и нереста лососей в руслах низшего порядка. Существенный
вклад в повышение продуктивности камчатских водотоков дают жилые формы колюшек
и гольцов, а также хариус, жизненный цикл которых по большей части приурочен к ручьям и
малым рекам (Зюганов, 1991; Скопец, 1993).
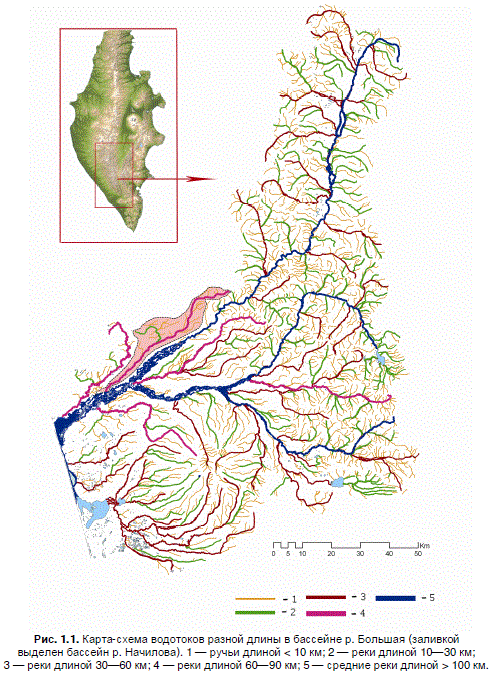
Рис. 1.1. Карта-схема водотоков разной длины в бассейне р. Большая (заливкой
выделен бассейн р. Начилова). 1 — ручьи длиной < 10 км; 2 — реки длиной 10—30 км;
3 — реки длиной 30—60 км; 4 — реки длиной 60—90 км; 5 — средние реки длиной > 100 км.
11
Таблица 1.3. Среднемноголетний потенциальный вклад (% от суммарной площади нерестилищ и абсолютные площади нерестилищ в га)
водотоков разной длины в воспроизводство стад тихоокеанских лососей бассейна р. Большая (по фондовым материалам КамчатНИРО)
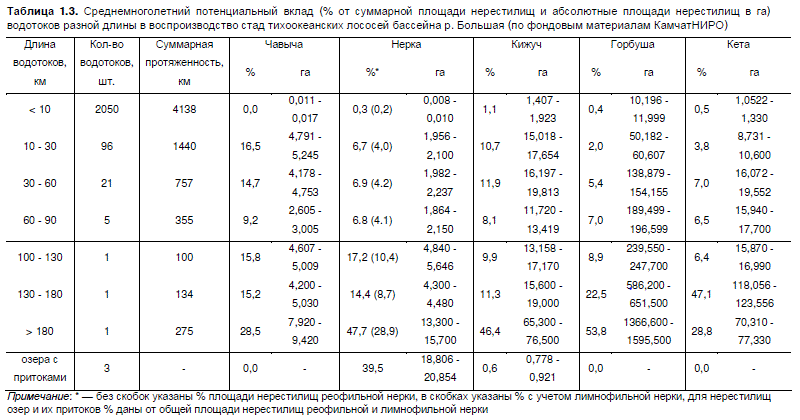
Примечание: * — без скобок указаны % площади нерестилищ реофильной нерки, в скобках указаны % с учетом лимнофильной нерки, для нерестилищ
озер и их притоков % даны от общей площади нерестилищ реофильной и лимнофильной нерки
Смотрите также
