Фенотипические различия молоди кижуча
Различия по скоростям роста
В мае 2007 г. на всем протяжении реки за исключением верховий с порожистыми рус-
лами ловились сеголетки кижуча длиной 27—31 мм, что соответствует размеру выхода личинок
кижуча из грунта (Смирнов, 1975; Зорбиди, 1988; Shapovalov, Taft, 1954). Выход молоди во всех
зонах продолжался до середины июля, при этом максимальная длина ранее вышедших сеголе-
ток к этому моменту была уже 65 мм (средняя июльская длина в бассейне в 2006 г. составила
40,0 мм). В сентябре средняя длина молоди составила 67,0 мм. Активный рост прекратился в
ноябре с понижением температуры воды до 2,0 (Бретт, 1983) — 2,5 ºС (McMahon, Hartman,
1989; Sandercock, 1991) и возобновился на следующий год в мае (температура воды в бассейне
10—11 мая была 2,8—3,5 ºС). Длина годовиков, которые ловились в начале мая, начиная с во-
дотоков с разветвленными горными руслами и ниже, колебалась в пределах 50—111 (в
среднем по бассейну 77,2) мм. Период активного роста в бассейне в 2006—2007 гг. составил,
таким образом, приблизительно 6 месяцев (см. рис. 2.2), различия его продолжительности
между верхним (станция Н3) и нижним (Н6) течением не превысили одной недели. К середине
июля второго года жизни 2006 г. кижуч вырос до 115—127 мм (средний бассейновый показатель
составил 93,2 мм), а к сентябрю до 142 (в среднем 121,6) мм (табл. 7.1).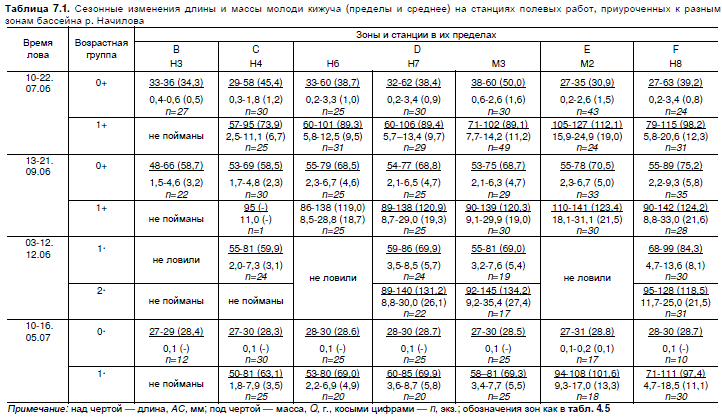
Примечание: над чертой — длина, АС, мм; под чертой — масса, Q, г., косыми цифрами — n, экз.; обозначения зон как в табл. 4.5
Для оценки групповой скорости роста в разных зонах использовали средние значения
длины молоди первого и второго года жизни, выловленной в бассейне в 2006—2007 гг.
(табл. 7.1). Было установлено, что в течение первого лета жизни наибольшей скоростью груп-
пового роста обладали сеголетки из уловов устьевой зоны (в среднем, 0,78 % длины тела в
сутки). Чуть медленнее росли сеголетки, обитающие на равнинных (в т.ч. "болотных") руслах
(0,72 %/сутки), причем если в июле-сентябре скорость роста на равнинных руслах с перекатами
не превышала 0,92 %/сутки, то на "болотном" участке она достигала 1,36 %/сутки. По-видимому,
различия в динамике летней скорости роста между этими зонами связаны с интенсивностью
прогрева воды в летне-осенний период (Corey et al., 1981). Так, на равнинных руслах с перека-
тами температура в июле колебалась от 11,8 до 12,4 ºС (среднесуточное значение 12,1 ºС), а
на "болотном" участке — от 13,1 до 16,0 (в среднем 13,9) ºС (см. табл. 4.8). В предгорном поясе
летом сеголетки росли медленнее: 0,59 %/сутки на неразветвленных и 0,61 %/сутки на разветв-
ленных руслах. Вода прогревалась здесь до 10,0 (в среднем 8,9) и 10,7 (9,1) ºС соответственно.
С октября по конец мая молодь кижуча в бассейне практически не росла, предельные скорости
достигали 0,1 %/сутки. Самый короткий период задержки роста зафиксирован на устьевом уча-
стке, где до декабря сохранялась скорость роста 0,14 %/сутки, самая высокая скорость роста в
период с декабря по май зафиксирована у молоди с разветвленных горных русел
(0,04 %/сутки). Очевидно, что специфика зимнего роста в этих речных зонах связана с
наличием непромерзающих местообитаний. На второй год жизни максимальной скоростью
роста обладала молодь с равнинных русел (в среднем 0,71—0,73 % длины тела в сутки), на
"болотном" и устьевом участках этот показатель был в 4 раза ниже (табл. 7.2).
Таблица 7.2. Изменение средней групповой скорости роста (% длины тела/сутки) молоди
кижуча из бассейна р. Начилова в течение июля 2006 — мая 2007 г.
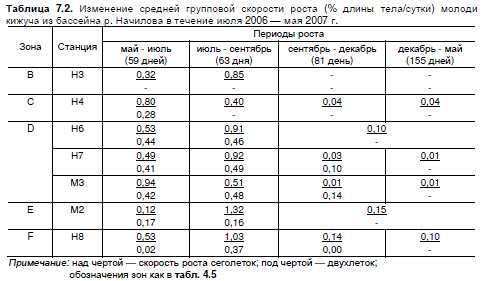
Примечание: над чертой — скорость роста сеголеток; под чертой — двухлеток;
обозначения зон как в табл. 4.5
В результате, к июлю второго года жизни наиболее мелкая молодь присутствовала в
уловах разветвленных горных русел, наиболее крупная — на "болотном" участке. Двухлетки
из водотоков с равнинными руслами обладали средними для бассейна размерами (табл. 7.1).
Распределения линейных размеров в выборках двухлеток из июльских уловов 2006 г. на
всех станциях, за исключением Н8, внешне носили симметричный одновершинный характер.
Асимметрия и эксцесс распределений в выборках из горных и равнинных русел были статисти-
чески не значимы. Нормальность распределения наблюденных значений свидетельствует о
морфологической гомогенности внутри выборок. Таким образом, в двухлетнем возрасте груп-
пировки кижуча бассейна р. Начилова не разделяются на мелких кочующих особей (floaters) и
крупных оседлых особей (residuals), которые выделяются у сеголеток в начале лета
(Кириллова, 2008; Chapman, 1966; Puckett, Dill, 1985; Martel, 1996). На устьевом участке
распределение линейных размеров было бимодальное. По-видимому, это связано присутст-
вием в уловах как местной молоди, так и иммигрантов из рр. Начилова и Большая. По критерию
113
Стьюдента молодь из разных зон достоверно различалась длиной во всех парах сравнения
(табл. 7.3), трансгрессия выборок не превышала 35 %.
Таблица 7.3. Оценки достоверности различий между длинами тела и приростами чешуи (мм)
кижуча 1+, выловленного в июле 2006 г. в разных зонах бассейна р. Начилова, по критерию
Стьюдента (сравнение с выборкой из зоны F выполнено формально, т.к. не соблюдено условие
нормальности ее распределения)
Смотрите также
