Эмбриональное развитие
Эмбриональное развитие красноперок изучалось нами в эксперименте и на естествен ных нерестилищах. Искусственно оплодотворенная (сухим способом) икра инкубирова лась в чашках Петри, помещенных в кристаллизаторы. Инкубация проводилась также ] садках из мельничного газа, на дно которых насыпалась галька, имитирующая нерести лище. Икра закапывалась в эту гальку, и садки помещались в ручей на течении. Парал лельно практически ежедневно брались пробы эмбрионов на нерестилищах. Производи тели Т. brandti для опытов вылавливались в Ударнице, Т. hakonensis и Т. ezoe — в озер* Русское. Рисунки, иллюстрирующие процесс развития, выполнены на живом материале.
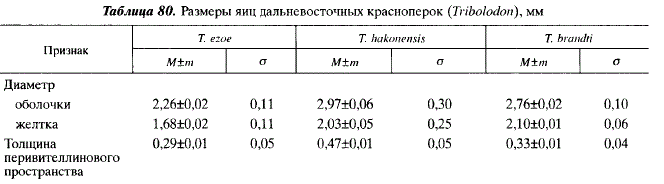
До попадания в воду зрелые ооциты имеют довольно прочные прозрачные оболочю толщиной 34-45 мкм, тесно прилегающие к плазматической мембране. В воде наружны] слой яйцевой оболочки, хорион, становится клейким, благодаря чему икринки способнь прикрепляться к субстрату. По данным наших наблюдений, яйца всех трех видов крас ноперок имеют клейкий хорион. Однако по данным М. Накамуры [Nakamura, 1969] 1 Ю.И. Гавренкова [1982], у отдельных популяций Т. hakonensis встречается неклейкая ик ра. Икра у всех видов красноперок окрашена в разные оттенки желто-оранжевого цвете Диаметр обводненных яиц разных видов красноперок различен, что может служит дополнительным таксономическим признаком (табл. 80). Использование этого признак может быть полезным при идентификации нерестилищ красноперок.
Все три вида красноперок имеют близкую продолжительность инкубационного периода (22-30 сут). Органогенез у этих видов в основном носит сходный характер (рис. 113-121, табл. 81-83). Различия между видами проявляются преимущественно в гетерохронности закладки тех или иных органов. Так, у Т. hakonensis несколько раньше, чем у Т. ezoe, происходит образование хвостовой почки и грудных плавников, черный пигмент на спине появляется на 4 сут раньше, чем у Т. ezoe. От двух других видов эмбрионы Т. hakonensis отличаются большим объемом желточного мешка в момент вылупления, более широкой хвостовой частью первично-плавниковой складки и большим диаметром глаза к моменту перехода в личиночный период развития. У Т. brandti раньше, чем у других видов, развивается перикардиальная полость, голова у вылупившихся эмбрионов загнута вниз в меньшей степени, чем у Т. hakonensis и Т. ezoe. Эмбрионы красноперок находятся в грунте нерестилищ практически до полной резорбции желтка.
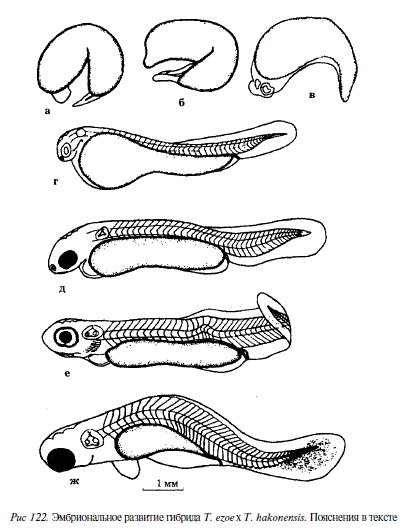
Одновременно с опытом по изучению нормального развития красноперок были поставлены опыты по реципрокному скрещиванию Т. ezoe и Т. hakonensis. Опыт с нормальным развитием Т. ezoe, описанный в табл. 81 и на рис. 113-115, использовался в качестве контрольного. В первом скрещивании икра одной самки Т. ezoe была оплодотворена молоками четырех самцов Т. hakonensis. Во втором скрещивании икра одной самки Т. hakonensis была оплодотворена молоками четырех самцов Т. ezoe. В последнем опыте не произошло набухания яиц и не наблюдалось их развития. Мы допускаем, что одной из причин этого было несовершенство проведения эксперимента. В первом опыте набухание произошло у всех яиц, однако объем перивителлинового пространства сильно варьировал. Дальнейшее развитие наблюдалось только у яиц с большим набуханием. До завершения гаструляции из 500 яиц дошло только 31. На 2-е сутки отчетливо сформировалось тело зародыша, началась сегментация туловищной мезодермы (4 сегмента). У многих зародышей не заложились глазные пузыри. Далее темп развития некоторых зародышей резко снизился. В возрасте 3 сут большинство из них имели недоразвитие головного отдела, у многих отсутствовали зачатки глаз (рис. 122, а, б, в). У отдельных яиц развитие задержалось на стадии мелкоклеточной морулы, но они не погибали в течение нескольких суток. На 5-е сутки оставшиеся в живых эмбрионы перешли в подвижное состояние. В возрасте 7,5 сут вылупились два эмбриона (см. рис. 122, г, д). На этом процесс вылупления завершился. Одна из этих двух предличинок имела нормальное строение, длина ее равнялась 5,5 мм; другая предличинка имела сильно искривленную хорду и тело. У большинства зародышей желточный мешок был аномальной формы. Начало пигментации глаз наблюдалось на 10-е сутки как у вылупившихся эмбрионов, так и у живых эмбрионов, не сумевших покинуть оболочку (см. рис. 122, е, ж). На этом эксперимент был прерван. Таким образом, гибридное потомство оказалось нежизнеспособным.
Аналогичные результаты были получены В.Н. Иванковым с соавторами (1987) при гибридизации Т. brandti и Т. hakonensis.
В литературе имеются сведения, что молодь красноперок задерживается в реках до 2 лет [Крыхтин, 1960; Казанский и др., 1968]. Специальные исследования, проведенные в ряде сахалинских рек (Богатая, Очепуха, Ударница, Первая Речка, Айруп), дают основание утверждать, что личинки всех трех видов красноперок скатываются из рек сразу же после выхода из грунта. Потомство раннерестующего вида Т. hakonensis, видимо, выносится остатками весеннего паводка. Личинки Т. ezoe в небольших количествах задерживаются на нагул в реках на несколько дней, но основная их масса после выхода из грунта транзитом проходит реку и попадает на нагул в море или в озера. Так, в течение августа мы наблюдали стайки личинок красноперок в низовье реки Богатая. Длина их колебалась от 9,5 до 14,7 мм, увеличения средней длины личинок в течение месяца не произошло. Личинки Т. brandti из реки Ударница сразу же после выхода из грунта попадают в озеро Тунайча.
Основанием для утверждения того, что молодь красноперки задерживается в реках до 2 лет, видимо, послужили случаи поимок в низовьях рек 2-леток, зашедших туда из моря. Нами подобные особи ловились в низовьях Танги и Первой Речки, однако численность их была крайне низка, значительно ниже обычно существующей в подобных реках численности производителей. Вместе с тем, мы наблюдали летний заход молоди красноперок из моря в низовье Богатой, где она до того времени отсутствовала. Мы допускаем, однако, возможность задержки молоди на год в крупных реках, таких как Тымь и Поронай.
Таблица 81. Эмбриональное развитие Tribolodon ezoe
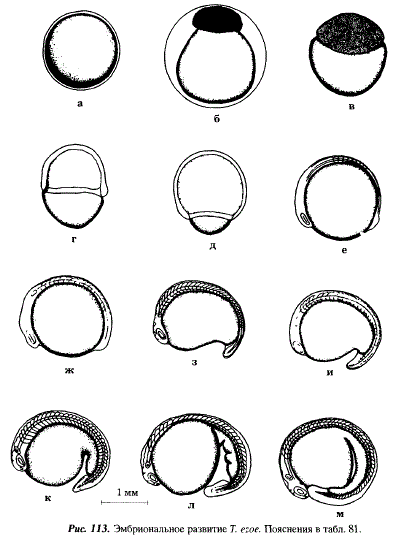
0 - Оплодотворение (см. рис.113,а)
1 - Набухание закончилось, на анимальном полюсе сформировался плазменный бугорок
2 - 2-4 бластомера
4 - 16-32 бластомера, начало крупноклеточной морулы
10 - Мелкоклеточная морула, бластомеры сгруппированы в форме шапки, контур ее компактнее, чем у других видов (см. рис.113,6)
16 - Начало гаструляции и эпиболии, бластодерма занимает около 30 % поверхности желтка, одновременно край бластодиска утолщается, образуется зародышевое кольцо (см. рис.113,в)
28 - Слой бластодермы охватывает более 50 % площади желтка, яйцо слегка вытянуто в анимально-вегетативном направлении (см. рис.113, г)
30 - После полного замыкания "желточной пробки" в результате конвергентных миграций глубоких клеток дифференцируется зародышевый щиток (см. рис. 113, д)
36 - В теле зародыша выделилось 7-8 пар сомитов, дифференцирован зачаток хорды, дифференцировались зачатки глазных пузырей (см. рис.113,е)
51 - В теле зародыша выделилось 15-17 пар сомитов, заложились слуховые плакоды, тело зародыша охватывает 2/3 желтка (см. рис.113,ж)
60 - Начала обособляться хвостовая почка, начало подвижного состояния зародыша, желточный мешок приобретает грушевидную форму с сужением в каудальном направлении, число пар сомитов возросло до 32-35 (см. рис.113,з,и)
72 - У зародыша в теле насчитывается до 37 миомеров, становятся заметны обонятельные плакоды, желток приобретает форму запятой, слуховые плакоды преобразовались в слуховые пузырьки (см. рис.113,к)
90 - В теле эмбрионов выделяется 39-40 мускульных сегментов, спинная часть непарной плавниковой складки доходит до 19-20-го туловищного сегмента, хорошо дифференцирована ее хвостовая часть, в глазных бокалах дифференцировались хрусталики (см. рис.113,л)
108 - Начало пульсировать сердце, оно представляет собой изогнутую трубку, окруженную перикардиальной полостью (см. рис.113,м)
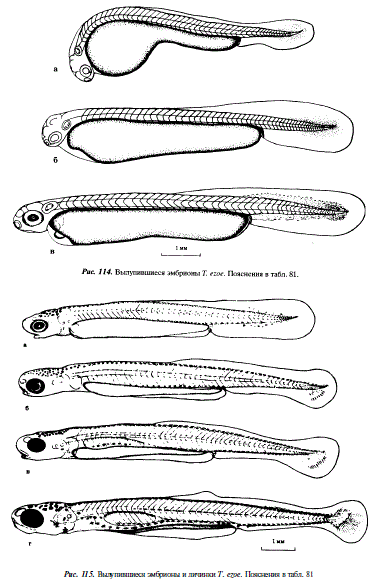
154 - Началось вылупление эмбрионов из яйцевых оболочек, длина эмбрионов 5,6-6,5 мм, в туловищном отделе 33-34 сегмента, в хвостовом 14-16, зачатки глаз не пигментированы, длина хвостового отдела составляет 25 % длины тела, спинной отдел непарной плавниковой складки начинается на уровне 12-го туловщного сегмента, в головном мозге четко дифференцированы три первичные отдела: передний, средний и задний; в слуховых пузырьках началось формирование отолитов, расстояние между ними и глазом равно ширине 2-3 туловищных сегментов (см. рис.114,а)
252 - На третьи сутки после вылупления сегментация тела полностью завершается, в туловищном отделе имеется до 75, а в хвостовом до 17 сегментов, заметно увеличение перикардиальной полости в передней части желточного мешка, эритроциты в массе имеют розовый цвет, в хвосте происходит концентрация мезенхимы, длина эмбрионов 6,3-8,0 мм (см. рис. 114,б)
288 - Начало пигментации ретины глаз, формируется преанальная часть плавниковой складки, рот нижний, челюстной аппарат неподвижный, слуховые капсулы максимально приближены к глазам, эмбрионы обладают отрицательным фототаксисом, их длина 6,5-8,4 мм (см. рис. 114,в)
324 - Появляются зачатки грудных плавников, пигментация тела отсутствует
432 - У свободных эмбрионов на голове, вдоль средней линии тела, на дорсальной стороне желточного мешка, появляются меланофоры, хорошо заметны лучи в хвостовой части непарной плавниковой складки, длина эмбрионов 7,3-9,0 мм
456 - Грудные плавники хорошо развиты, рот из нижнего превращается в полунижний, появляется пигмент в хвостовом отделе и на нижних окончаниях миомеров (нижне-боковой пигментный ряд), (см. рис. 115,а)
576 - Длина эмбрионов 9,9-11,3 мм, желточный мешок сильно резорбирован, заметно увеличилась ширина преанальной части непарной плавниковой складки, ротовой аппарат стал подвижным (см. рис. 115,6)
600 - На дорсальной стороне тела эмбрионов появился желтый пигмент, имеется зачаток плавательного пузыря, дорсальная часть которого покрыта меланофорами (см. рис. 115,в)
720 - Длина эмбрионов 9,7-11,5 мм, плавательный пузырь наполнен воздухом, но еще не разделен на отделы, желточный мешок представляет собой узкий тяж, лежащий выше обособившейся преанальной части непарной плавниковой складки, сформированы жаберные крышки, на которых заметны крупные звездчатые меланофоры, в плавниковой складке появились расширения — будущие спинной и анальный плавники, в хвостовом плавнике появились гипуралии, образовавшиеся из задних хвостовых сегментов, глаза окрашены в черный цвет, их диаметр равен ширине четырех туловищных сегментов; эмбрионы положительно реагируют на свет, всплывают на поверхность, в потоке ориентированы против течения, переходят на внешнее питание, вступают в личиночный период развития (см. рис.115,г)
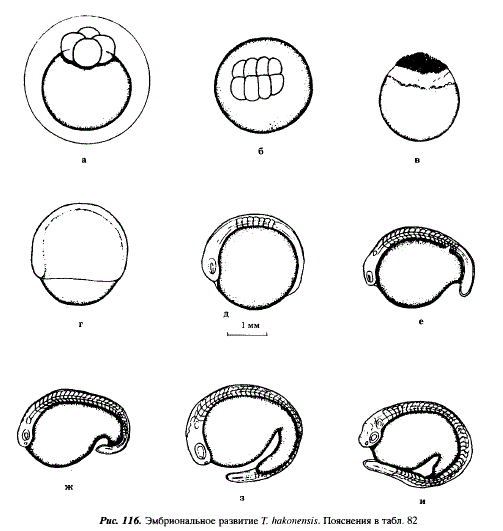
Таблица 82. Эмбриональное развитие Tribolodon hakonensis
0 - Оплодотворение
3 - Стадия четырех бластомеров (см. рис. 116,а)
4 - Стадия 8-16 бластомеров (см. рис. 116,6)
14 - Мелкоклеточная морула, вокруг основания которой имеется белесое кольцо, опоясывающее бластодиск (перибласт) (см. рис. 116,в)
28 - Начало гаструляции
36 - Бластодерма охватывает около 70 % поверхности желтка, происходит образование зародышевого щитка на дорсальной стороне желтка (см. рис. 116,г)
48 - Дифференцировался зачаток хорды, в теле зародыша выделилось 7 пар сомитов, сформировались глазные пузыри (см. рис. 116,д)
60 - В теле зародыша выделилось 18 пар сомитов, дифференцировались слуховые плакоды, начала обособляться хвостовая почка, желток приобретает грушевидную форму, начинается преобразование глазных пузырей в глазные бокалы (см. рис. 116,е)
72 - В теле зародыша выделяется 28 пар сомитов, появились обонятельные плакоды, дифференцировались три первичных отдела головного мозга (см. рис. 116,ж)
96 - Начало подвижного состояния зародыша, количество сегментов в теле зародыша увеличилось до 44 (см. рис. 116,з)
120 - Эмбрионы готовы к вылуплению, высокоактивны, сердце пульсирует, в теле насчитывается 45 сегментов, слуховые пузырьки преобразовались в слуховые капсулы, хорошо развит хвост (см. рис. 116,и)
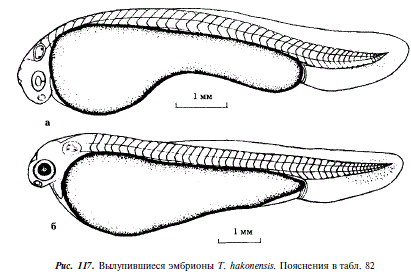
180 - Началось вылупление, эмбрионы имеют массивный желточный мешок слабо выраженной грушевидной формы, хвостовая часть непарной плавниковой складки более широкая, чем у Т. ezoe, хвостовой отдел относительно короткий, в слуховых капсулах образовались отолиты, в туловищном отделе 32, в хвостовом до 16 сегментов, длина эмбрионов 5,9-6,7 мм (см. рис. 117,а)
252 - У свободного эмбриона имеется хорошо выраженная перикардиальная полость, в сердце различимы желудочки и предсердие, кровь бесцветна, глаза пигментированы (см. рис. 117,б)
312 - Дифференцировались грудные плавники
336 - На дорсальной стороне тела появились меланофоры
480 - Длина свободных эмбрионов 12,4 мм
518 - В хвостовой части непарной плавниковой складки хорошо заметны лучи
600 - Плавательный пузырь наполнился воздухом
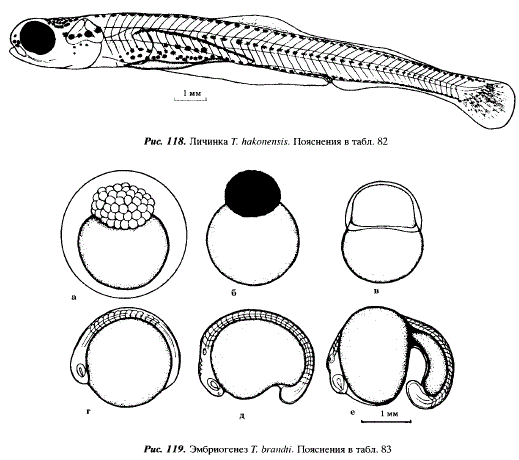
720 - Длина эмбрионов 13,4 мм, на дорсальной стороне тела имеется желтый пигмент, образовались гипуралии, расстояние между глазом и слуховой капсулой равно ширине 1,0-1,5 туловищных сегментов, наибольший диаметр глаза равен ширине 4—5 туловищных сегментов, глаза окрашены в черный цвет; эмбрионы переходят к личиночному периоду жизни, не прячутся от света, всплывают к поверхности воды, переходят на внешнее питание (см. рис. 118)
Таблица 83. Эмбриональное развитие Tribolodon brandti
0 - Оплодотворение
3,5 Стадия 2-4 бластомеров
8 Крупноклеточная морула (см. рис. 119,а)
18 Мелкоклеточная морула (см. рис. 119,6)
30 Эпиболия. Под бластодермой скрыто около 45 % поверхности желтка (см. рис. 119,и)
48 Начало гаструляции
72 Формируется зародыш
84 Головной отдел расширен, хорошо видны глазные пузыри, хорда хорошо обособлена, в теле зародыша выделилось 10-13 пар сомитов (см. рис.119,г)
108 Начало подвижного состояния эмбриона, начала обособляться хвостовая почка, дифференцировались слуховые и обонятельные плакоды, намечается разделение головного мозга на первичные отделы, желток приобретает грушевидную форму, количество мускульных сегментов достигает 30 (см. рис. 119,д)
134 Число мускульных сегментов увеличилось до 45-47, все зародыши имеют желточный мешок в виде запятой, перед желточным мешком под головным отделом образовалась перикардиальная полость, глазные пузыри преобразовались в глазные бокалы, в которых дифференцировались хрусталики, расстояние между слуховыми пузырями и глазом равно ширине трех туловищных сегментов (см. рис. 119,е)
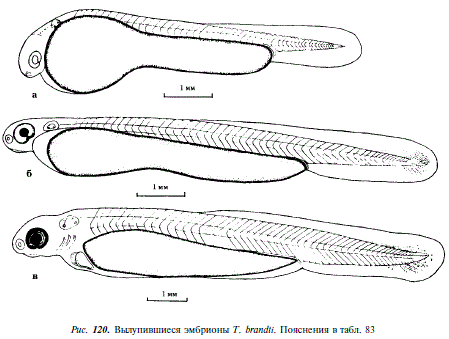
156 Начало вылупления; длина вылупившихся эмбрионов 6,6-7,2 мм, сердце пульсирует, функционирующих кровеносных сосудов обнаружить не удается, перикардиальная полость увеличилась, хвостовая часть непарной плавниковой складки хорошо развита, ее ширина равна ширине 7 туловищных сегментов, спинная часть ее берет начало от задней границы 20-го туловищного сегмента и, постепенно возвышаясь, окаймляет хвостовой отдел, заканчиваясь у анального отверстия, голова эмбриона загнута вниз, в слуховых пузырях происходит закладка отолитов (см. рис. 120,а)
216 Через двое суток после вылупления в ретине глаз эмбрионов появляется черный пигмент, в хвостовом отделе концентрируется мезенхима, клетки крови в массе окрашены в розовый цвет, продолжает увеличиваться объем перикардиальной полости, длина эмбриона 9-10 мм (см. рис. 120,6)
300 Формируется преанальная часть непарной плавниковой складки, глаза сильно пигментированы, расстояние между глазом и слуховым пузырем сократилось до ширины 1,5-2,0 туловищных сегментов, пигментация тела еще не началась (см. рис. 120,в)
336 Появились меланофоры на голове и на дорсальной стороне тела, появились зачатки грудных плавников
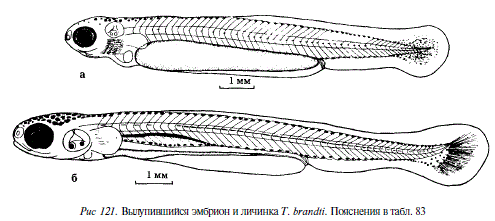
360 Появились меланофоры вдоль средней линии тела, в нижнебоковом и подхвостовом рядах, длина эмбрионов 10,5-11,8 мм, расстояние между глазом и слуховым пузырем сократилось до ширины 1,0-1,5 туловищных сегментов, в слуховых пузырьках образовались полуокружные каналы, за глазом видны четыре жаберных дуги со сформировавшимися на них жаберными лепестками, дуги не прикрыты жаберными крышками, преанальная часть плавниковой складки и рот хорошо дифференцированы, плавательный пузырь не наполнен воздухом, в хвостовой части протоптеригиальной складки начали формироваться плавниковые лучи (см. рис. 121,а)
480 Плавательный пузырь содержит воздух, слегка намечаются зачатки спинного и анального плавников, жабры прикртыты крышками, в хвостовом и грудных плавниках образовались лучи, появились гипуралии, длина эмбрионов 12,5-12,7 мм
528 Длина личинки 12,6-12,8 мм, объем воздуха, содержащегося в плавательном пузыре увеличился, ротовой аппарат приобрел подвижность, принял конечное положение, желток рассосался почти полностью, грудные плавники большие, расположенные вертикально, диаметр глаза равен ширине четырех туловищных сегментов, хорошо развита пигментация на голове и спине, плавательный пузырь сверху густо покрыт меланофорами, личинки активно плавают, переходят на внешнее питание (см. рис. 121,6)
768 В возрасте 32 суток плавательный пузырь у личинки разделен, рот типично передний, происходит обособление спинного плавника, в котором заметно 5 лучей, в хвостовом плавнике наметилась выемка, обосабливается анальный плавник, личинки плавают и питаются