Тихоокеанская (зубастая) корюшка
Тихоокеанская (зубастая) корюшка, обитающая в бассейне Тихого океана и арктических морей, рассматривается, согласно последней ревизии [Клюканов, 1969], в качестве подвида Osmerus mordax (Mitchill), которому дано латинское название Osmerus mordax dentex Steindachner. Мы разделяем точку зрения В.А Клюканова относительно таксономического статуса зубастой корюшки Дальнего Востока.
На Сахалине этот вид заходит для нереста в реки по всему побережью за исключением участка от мыса Терпения до Луньского залива, где береговая линия сильно спрямлена и сказывается влияние холодного Сахалинского течения. Отсутствует зубастая корюшка также в бассейне озера Тунайча.
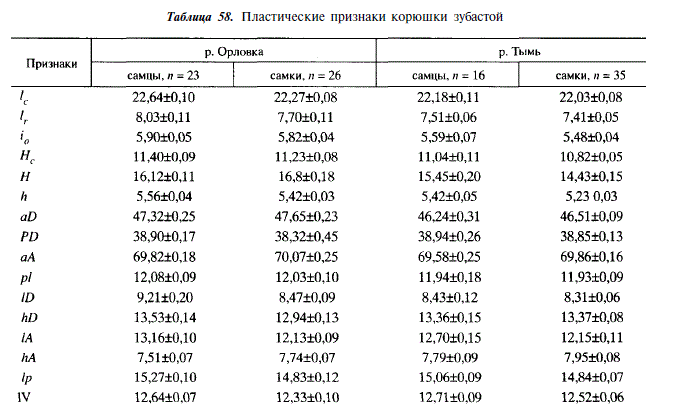
Подробное морфометрическое описание корюшки было сделано нами для ряда популяций острова Сахалин и для корюшки реки Камчатка. В данной работе мы приводим меристические признаки всех исследованных популяций, а пластические — только для рыб рек Тымь и Орловка. Последнее делается с целью сравнения пропорций тела у особей из большой и малой рек, относящихся к тому же к бассейнам различных морей. Как следует из табл. 56-58, зубастая корюшка в пределах Сахалина характеризуется значительным фенотипическим однообразием. Подобное однообразие, видимо, свойственно ей на большей части дальневосточного региона, так как сахалинские популяции фенотипически весьма сходны с камчатской. Попарное сравнивание признаков, приведенных в табл. 57, показывает, что каждая популяция достоверно отличается от остальных как минимум по одному признаку, а чаще — по нескольким. Величина этих различий далеко не достигает подвидового уровня (CD 1,28). Исключение составляет число жаберных лучей у корюшки из реки Гастеловка (при сравнении с другими популяциями CD оказывается больше 1,28). Величина различий между камчатской корюшкой и сахалинской не больше различий, существующих в пределах Сахалина. Различия по пластическим признакам также незначительны. Корюшка из Тыми отличается от корюшки из Орловки несколько меньшей головой, более низкими хвостовым стеблем, спинным плавником, расположенным ближе к голове, меньшей высотой тела (однако последнее, возможно, объясняется различной зрелостью гонад в период взятия проб).
По характеру нереста зубастая корюшка в реках Сахалина является типичным литофилом, в отличие от обской, откладывающей икру на прогруженную в воду или свободно плавающую травянистую и древесную растительность [Амстиславский, 1959; Венглинский и др., 1967]. Нерестится она на каменисто-галечных перекатах со скоростью течения 0,6-0,9 м/с. Глубина в местах нереста 0,2-0,5 м. Во время паводков глубина и скорость течения увеличиваются. В различных реках острова места, пригодные для нереста корюшки, находятся на разном удалении от устьев, что во многом определяет особенности нерестовой миграции и нереста [Гриценко и др., 19846].
У сахалинской корюшки выделяются три группы популяций (три экотипа), различающихся по комплексу признаков, связанных с размножением. Популяции первой группы воспроизводятся в малых реках, где нерестилища расположены непосредственно выше устья и зоны подпора; протяженность их обычно не более нескольких сотен метров. Производители заходят в реку только на ночные часы и по совершении акта нереста скатываются под утро в море. Нерестовый период длится около 1,0-1,5 мес, начинается после
прохождения весеннего паводка и заканчивается в период летней межени. Температура воды во время нереста обычно колеблется от 10 до 14°С. Подобная ситуация наблюдается в реках Очепуха, Вишера, Первая Речка.
Популяции второй группы воспроизводятся в малых и средних реках. Нерестилища в них начинаются на расстоянии от 1-2 км от устья и занимают участок русла длиной от 1 до 5 км. Производители на нерестилищах находятся круглосуточно, постепенно заменяясь особями новых подходов. Нерест длится 1,0-1,5 мес. Начинается он после прохождения пика весеннего паводка, а заканчивается в период летней межени. Подобная картина наблюдается на реках Хоэ, Урюм, Фирсовка.
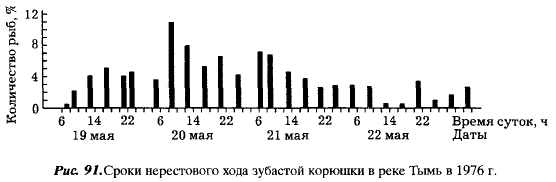
Популяции третьей группы воспроизводятся в наиболее крупных реках — Тыми и Поронае, где нерестилища удалены от устья на несколько десятков километров. В Тымь корюшка начинает заходить во П-Ш декаде мая, через 2-3 дня после окончания ледохода, при температуре воды 2-3 °С. Нерестовая миграция приходится на время весеннего паводка. Нерестовый ход и нерест сравнительно короткие. Вся масса производителей заходит в реку в течение нескольких дней и непрерывным потоком движется к нерестилищам, расположенным в 55-70 км от устья, где находятся первые галечные перекаты с небольшими глубинами (рис. 91). Путь до нерестилищ занимает около 4 сут. По мере подъема производителей по реке вода в ней прогревается до 6-7 °С и вскоре после нереста повышается до 9-17 °С. Нерест длится 6-9 дней и приходится на высокий уровень паводка, после чего производители скатываются из рек (рис. 92).
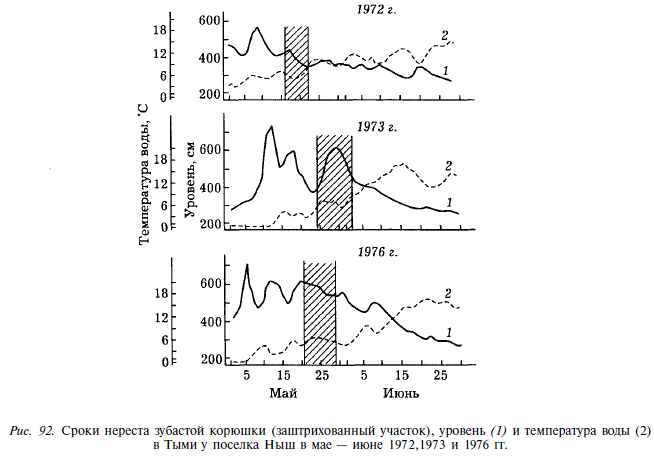
Таким образом, в реках, различающихся по протяженности и широтному расположению, нерест и эмбриогенез зубастой корюшки происходят практически при одной и той же температуре воды (10-14°С). Различия сроков захода производителей в реки, так же как и различия гидрологических условий в начале нерестовой миграции, связаны с необходимостью достичь нерестилищ к моменту прогрева воды до температуры, при которой происходит нерест.
В период нерестовой миграции и нереста корюшка в реках Сахалина не питается. Прекращение питания в этот период отмечено для корюшки Амура и Оби [Никольский, 1956 б; Амстиславский, Брусынина, 1963]. В Енисее, где нерестилища расположены на расстоянии свыше 100 км от устья, корюшка продолжает питаться во время нерестовой миграции [Тюрин, 1924].
Продолжительность инкубационного периода определялась нами экспериментально. Икра, оплодотворенная сухим способом, инкубировалась в чашках Петри при температуре 4,6-17,8 °С. Вылупление началось в возрасте 18 сут при 172 градусо-днях, массовое вылупление наблюдалось в возрасте 24 сут при 272 градусо-днях, закончилось вылупление на 25-е сутки при 284 градусо-днях. Длина предличинок в момент вылупления 7-8 мм. Наши данные о числе градусо-дней сходны с результатами Янагавы [Yanagawa, 1978].
Фрагментарные наблюдения за скатом личинок корюшки проведены на реке Урюм 10 и 11 июня, — через 27 дней после начала массового нереста, конической ловушкой из мельничного газа, выставляемой ниже нерестилищ; собраны пробы сносимых течением икринок и личинок. В уловах были вылупившиеся предличинки и живые икринки корюшки; кроме того, было отмечено большое количество мертвой икры и пустых оболочек. Предличинки составляли 29 % от общего числа живых предличинок и икринок; икринки с зародышами, имеющими пигментированные глаза, — 42%, непигментированные глаза — 29 %. Длина предличинок 5,2-7,8 мм.
Поскольку в большинстве сахалинских рек температура воды в период инкубации икры корюшки колеблется от 9 до 18 °С, развитие ее должно продолжаться 20-30 дней. Следовательно, основная масса личинок попадает в море в течение июня и первой декады июля. Заканчиваться скат должен в последних числах июля. Естественно, часть икринок, оторвавшихся от субстрата, в коротких реках выносится в море ранее этих сроков, что возможно, вызывает их гибель.
Скат корюшки на Сахалине значительно отличается от ее ската на Амуре. Ю.Н. Подушко [1970а], изучавшая скат в устье Амура в районе города Николаевска, не встречала сносимых течением икринок, в уловах были только предличинки, имевшие относительно большой желточный мешок.
В нерестовых стадах корюшки заметно преобладают самцы. Эта особенность объясняется ранним созреванием самцов и, соответственно, меньшей элиминацией, чем у самок, к моменту нереста, а также более длительным пребыванием самцов на нерестилищах. В отдельных случаях доля самцов на нерестилищах достигает 80-90 % [Гриценко и др., 19846].
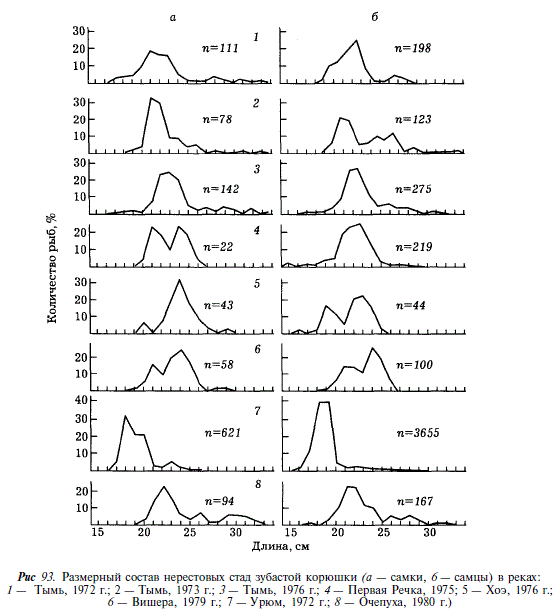
Размеры производителей зубастой корюшки колеблются от 15 до 34 см. Особи длиной 21-24 см составляют примерно 80 %. Самцы несколько мельче самок. Как показано на рис. 93, в реках Сахалина нерестовые стада корюшки мало различаются по размерному составу. В нашем материале несколько особняком стоит корюшка из реки Урюм, выделяющаяся малыми размерами. Доля крупных рыб (более 25 см) в Тыми больше, чем в малых реках.
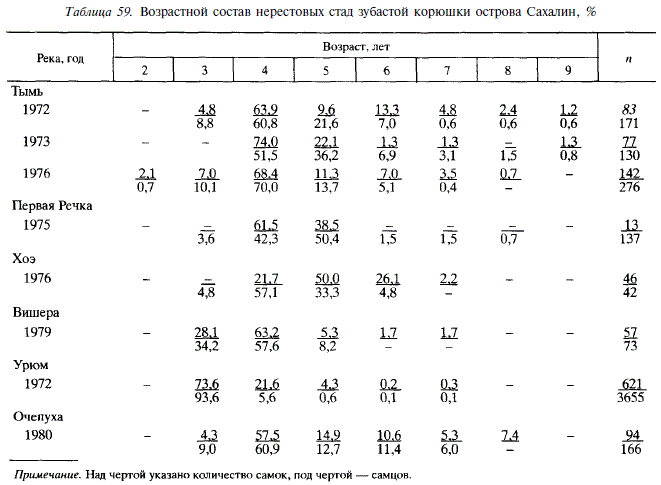
Нерестовые стада корюшки состоят из особей в возрасте от 2 до 9 лет, однако 90 % рыб приходится на 3-5-годовиков при резком преобладании 4-годовиков (табл. 59). Эта возрастная группа доминировала как в Тыми в течение всех 3 лет наблюдений, так и в малых реках (Очепуха, Вишера, Первая Речка). Корюшка из реки Урюм выделяется более ранним созреванием (в основном 3 года). Доля рыб старших возрастов в реке Тымь выше, чем в других реках. Нерест у корюшки неоднократный, однако большинство особей до второго нереста не доживают. Сахалинская корюшка крупнее, чем амурская, и созревает в массе на год позже амурской [Подушко, 19706; Гриценко и др., 1984а]. Корюшка из реки Урюм по размерному и возрастному составу весьма напоминает амурскую.
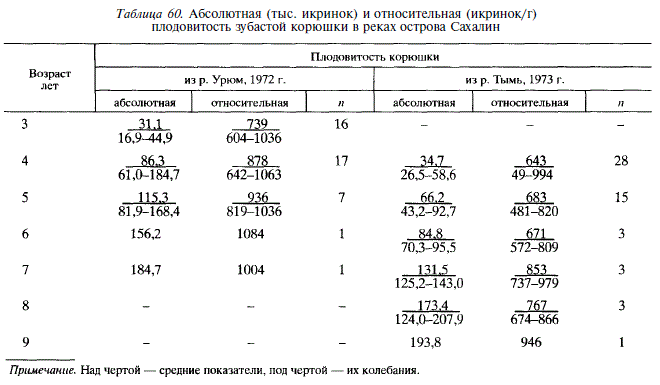
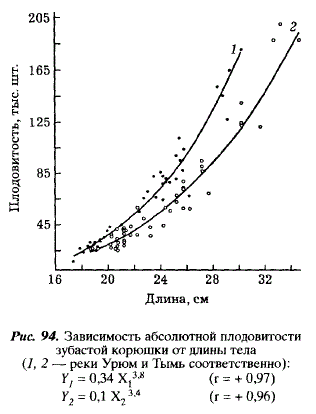
В водах Сахалина индивидуальная абсолютная плодовитость корюшки колеблется от 16,9 до 207,9 тыс. икринок. Сравнение корюшки из рек Урюм и Тымь обнаруживает существенные популяционные различия: в Урюме плодовитость одновозрастных рыб выше, чем в Тыми (табл. 60); выше в Урюме и плодовитость одноразмерных особей (рис. 94).
Большая по сравнению с тымской плодовитость корюшки из реки Урюм наряду с различиями длины рыб свидетельствует о различном соотношении пластического и генеративного обмена у этих двух стад и, естественно, о различиях отношений со средой. Видимо, в малых реках смертность корюшки на ранних стадиях развития выше, чем в больших, из-за раннего выноса в море личинок и развивающихся икринок, что и выражено как адаптивный ответ в виде большей плодовитости корюшки в малых реках. Наше предположение подтверждается тем, что плодовитость амурской корюшки [Подушко, 1971] гораздо ниже, чем корюшки из Урюма, и близка к плодовитости корюшки из Тыми.
Отнерестовавшие производители корюшки появляются в Ныйском заливе с конца мая и в течение лета и осени постоянно обитают в нем. Зимовка происходит как в типично морских условиях, так и в солоноватоводных заливах.
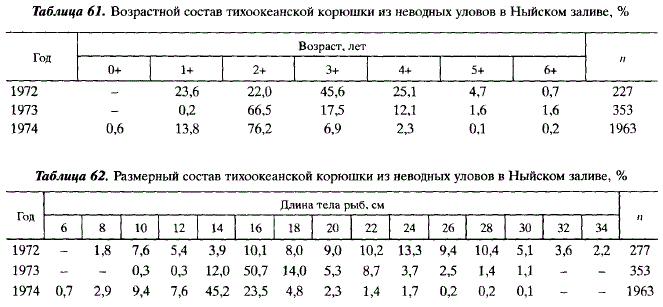
В нагульный период корюшка отлавливалась нами мелкоячейным неводом и жаберными сетями. Неводные уловы представительно характеризовали возрастной состав относительно молодых особей (Он—6+ (табл. 61), сетные уловы — относительно старых (4+-9+), составляющих нисходящую часть кривых размерного и возрастного состава.
В Ныйском заливе в уловах встречались особи длиной от 4, 5 до 35,5 см (табл. 62) и массой от 0,4 до 357 г. В неводных уловах в 1973-1974 гг. преобладали 3и 4-летки — в 1972 г. (см. табл. 61). Сеголетки отмечены в небольшом количестве лишь в 1974 г. Скорее всего, сеголетки и 2-летки нагуливаются вне залива, иначе при малых глубинах залива их доля в уловах должна была бы быть значительной.
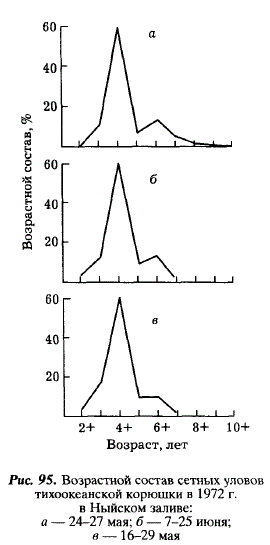
Анализируя сетные уловы, мы видим, что к июню из них исчезают неоднократно нерестовавшие старшие рыбы (7+-9+) (рис. 95). Их элиминация, видимо, объясняется посленерестовой гибелью. Из рис. 96 видно, что эти рыбы начинают питаться гораздо позже мелких особей. Средний улов корюшки на одну постановку сети снижается со 161 экз. в конце мая до 121 экз. в июне и 80 экз. во второй половине июля. Не исключая возможности миграции части рыб за пределы залива, мы склонны ведущую роль подобного уменьшения отвести смертности, поскольку в нерестовом стаде корюшки из Тыми доминируют 4-годовики, а доля 5-годовиков резко снижается. Основными факторами смертности, наиболее вероятно, являются изменения физиологического состояния производителей после нереста и выедание хищными рыбами, в частности, сахалинским тайменем [Гриценко, Чуриков, 19776]. 5-годовики длиной 22 см, составляющие модальную размерную группу в неводных уловах, являются также модальной группой корюшки, извлеченной из желудков тайменя.
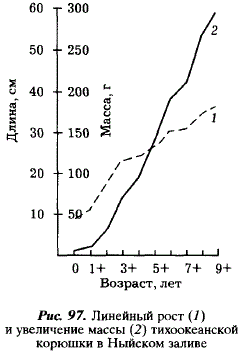
В линейном росте корюшки выделяется два периода: ускоренный (0+-3+) и относительно замедленный (4+ -9+), наблюдаемый у рыб, достигших половозрелости. Рост массы ускоряется на 3-м году и до конца жизни остается относительно постоянным (см. рис. 96).
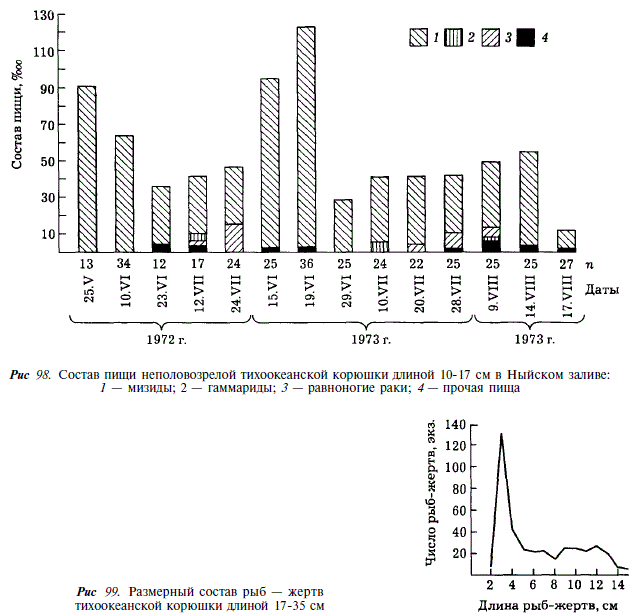
Основу рациона неполовозрелой корюшки длиной 10-17 см составляют ракообразные, среди которых преобладают мизиды. На долю рыбной пищи неполовозрелой корюшки приходится около 4 % (рис. 97). Состав пищи остается относительно постоянным в разные годы. Наибольшие индексы наполнения желудков отмечаются а мае — июне, в начале нагульного периода [Чуриков, Гриценко, 1983].
Пищевой рацион половозрелой корюшки в Ныйском заливе состоит из нектобентических ракообразных и рыбы (рис. 98). Среди ракообразных преобладают мизиды, составляющие 30-50 % массы пищи. Рыбную часть пищи составляют 14 видов, относящихся к 8-ми семействам. Кроме того, корюшка потребляет молодь тихоокеанской проходной миноги. Основу рыбной пищи составляют 4 вида: сельдь, малоротная корюшка Н. olidus, мойва и навага, на долю которых приходится от 50 до 80 % всей съеденной рыбы. Видовой состав пищевых компонентов постоянен из года в год, соотношение же их значительно меняется. В течение летнего нагула наиболее низкое наполнение желудков у половозрелой корюшки наблюдается в первые дни после ската с нерестилищ. При этом у более крупных рыб наполнение ниже, чем у мелких. Спустя некоторое время после ската в залив, индексы наполнения возрастают. В отдельные периоды посленерестового нагула наблюдается резкая смена пищевых компонентов, обусловленная появлением в большом количестве тех или иных кормовых объектов (например, нерестовой мойвы или молоди сельди).
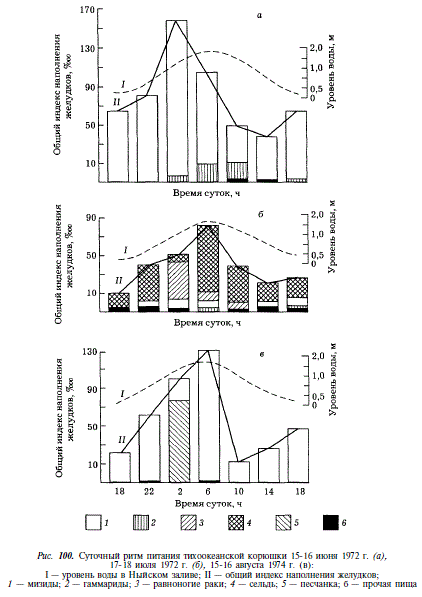
Подобно кундже, корюшка, потребляя рыб, выступает как макропланктофаг и как типичный хищник. В первом случае она кормится на массовых скоплениях мелких рыб длиной 2,5-5,0 см (колюшка девятииглая, песчанка, молодь кеты и горбуши, бельдюга), во втором случае она потребляет нерестовую мойву, малоротую корюшку, молодь сельди. Подобный характер питания наглядно демонстрируется размерным составом рыб-жертв, извлеченных из желудков корюшки (рис. 99).
Тихоокеанская корюшка является основным потребителем молоди тихоокеанских лососей в Ныйском заливе. Выедание ею покатной молоди кеты и горбуши происходит в период наиболее интенсивного ската и может рассматриваться как один из факторов, влияющих на формирование численности этих видов лососей [Чуриков, 1975а].
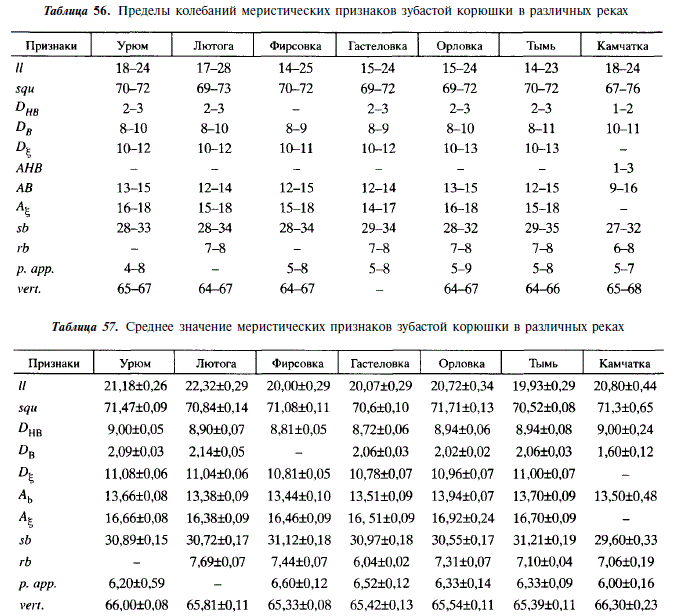
Суточный ритм питания корюшки имеет четко выраженный моноциклический характер (рис. 100).
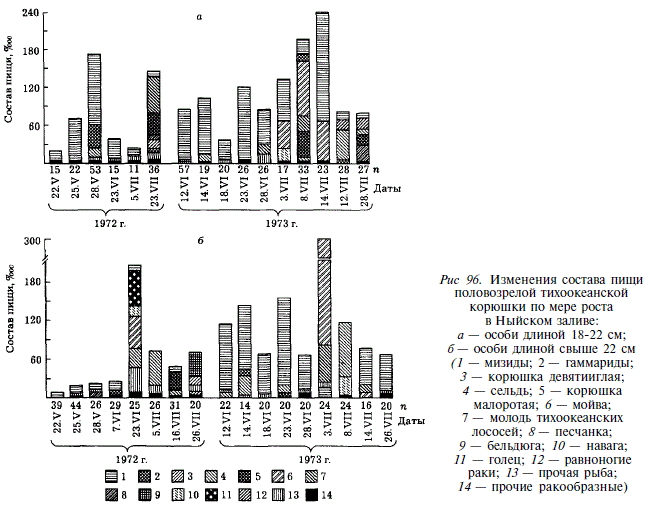
Интенсивность питания повышается в вечерние часы в начальную фазу прилива, достигает максимума ночью, а утром с отливом снижается. На ее характер, несомненно, оказывает влияние освещенность. Так, 13-16 июня, в дни, близкие к летнему солнцестоянию, максимальная наполненность желудков отмечалась в 2ч; 17-18 июля и 15-16 августа, с увеличением продолжительности ночи, она стала наблюдаться в 6 ч. Состав пищи в течение суточного цикла практически не меняется: 15-16 июня и 15-16 августа потреблялись преимущественно мизиды, 17-18 июля — сельдь.
Исходя из общих закономерностей трансформации энергии по пищевым цепям от продуцентов к консументам через ряд трофических уровней [Одум, 1975] и проведенного анализа питания корюшки, можно заключить, что ее биомасса создается по детритному и пастбищному каналам (пищевым цепям) при значительном преобладании первого. Действительно, основу питания тихоокеанской корюшки составляют ракообразные-детритофаги и рыбы-нектобентофаги, поедающие этих ракообразных, за счет этих двух компонентов создается основная часть ее биомассы. Пастбищный канал корюшка использует, потребляя сельдь и мойву, продукция которых создается вне залива за счет потребления зоопланктона открытого моря.