Род таймени Hucho Gunther
До недавнего времени считалось, что род Hucho представлен пятью видами: Н. hucho (L.) — дунайский лосось (бассейн Дуная), Н. taimen (Pallas) — сибирский таймень (реки Сибири, а также бассейны рек Печора и Кама), Н. perryi (Brevoort) — сахалинский таймень (реки Приморья, Сахалина, Хоккайдо и северной части Хонсю), Н. ishikawai Mori (река Ялу, Корейский п-ов), Н. bleekeri Kimura (верховья Янцзы) [Берг, 1948; Nakamura, 1963]. Исследованиями последних двух десятилетий было показано, что различия, существующие между Н. hucho и Н. taimen, незначительны и соответствуют скорее подвидовому уровню. Практически различить их можно только по характеру брачной окраски [Шапошникова, 1968; Гриценко, Чуриков, 1980; Holcik, 1982].
Номенклатурное объединение этих двух видов было проведено Ю. Холчиком с сотрудниками [Holfrk, et al., 1984]. По приоритету дунайский лосось рассматривается в качестве номинативного подвида Н. hucho hucho, за сибирским тайменем закреплено название Н. hucho taimen. К подвиду Н. hucho hucho Ю. Холчик с сотрудниками относят только дунайского лосося.
Биология видов Н. ishikawai и Н. bleekeri практически не изучена, описаны они по единичным экземплярам, в связи с чем, нам кажется, их следует рассматривать как выделенные провизорно. Первый из них, по мнению Ю. Холчика и соавторов [Holcik, et al., 1984], возможно, является синонимом Н. hucho taimen, так как у обоих число прободенных чешуи в боковой линии меньше числа поперечных рядов чешуи. У сахалинского тайменя значения этих признаков практически одинаковы. В пользу реальности Н. bleekeri свидетельствует малое число пилорических придатков (табл. 50). К сожалению, в описании данного тайменя С. Кимура [Kimura, 1934] не приводит точного значения этого признака, указывая, что придатков более 60.

Существенной ревизии подвергся родовой статус тайменей. В. Владыков [Vladykov, 1963] на основании особенностей географического распространения и образа жизни и Г.Х. Шапошникова [1968] на основании остеологических различий между сахалинским тайменем и тайменями дунайским и сибирским выделяют первый вид в монотипический подрод Parahucho Vladykov. Этот подрод характеризуется проходным образом жизни, наличием костной пластины на basibranchialia, наличием мелких зубов посередине linguale, выступом внешнего края frontale над sphenoticum. Обоснованность подобного разделения принимается Ю. Холчиком [НоШк, 1982]. В.П. Васильев на основании различий кариотипов допускает вероятность принадлежности Я. penyi и Я. hucho к разным родам: у Я. hucho hucho 2 п = 82; NF = 106; у Я. h. taimen 2 п = 84; NF = 102; у Я. perryi 2 п = 62; NF = 100 [Васильев, 1985].
Подобное разделение рода тайменей не представляется обоснованным. Чисто субъективно перечисленные различия нам кажутся недостаточными для выхода за рамки известной осторожности, требуемой при выделении монотипических таксонов крупнее вида, ибо род тайменей практически воспринимается как целостное природное явление, сопоставимое с такими явлениями (и качественно отличное от них), как роды гольцов или тихоокеанских лососей. Более того, если Я. hucho и Я. taimen являются единым видом (как это считают Г.Х. Шапошникова и Ю. Холчик), то, сравнивая его с Я. perryi, следует говорить не об отдаленности последнего от "компактной группы видов", а лишь о дистанции между двумя видами, что существенно уменьшает основания для таксономического подразделения рода.
В вопросе о двух подвидах вида Я. hucho мы считаем необходимым сделать следующее уточнение. Поскольку основой для идентификации подвидов служит тип брачной окраски, таймень бассейна Камы должен быть отнесен по этому признаку к подвиду Я. hucho hucho, а не к Я. hucho taimen (рис. 86), в связи с чем следует говорить о европейском и сибирском подвидах Я. hucho, граница между которыми проходит по Уралу.

Попытка систематизировать имеющиеся в литературе сведения по нерестовой окраске тайменей показала, что в Сибири встречаются два типа окраски, не связанные с половым диморфизмом (см. рис. 86). Это позволяет предположить существование двух репродуктивно обособленных форм тайменя. Подобные различия в окраске могут быть использованы в качестве ориентира в дальнейших исследованиях систематики тайменей.
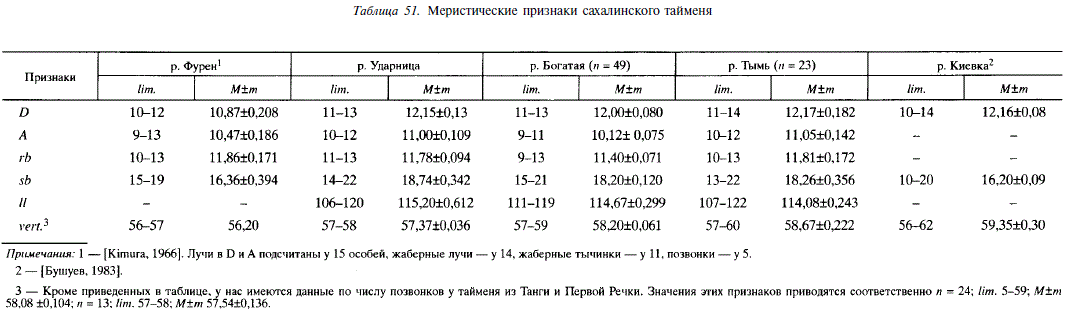
В пределах ареала сахалинский таймень предстает морфоэкологическим однородным видом. Межпопуляционные различия меристических признаков у него незначительны. Закономерные изменения отмечаются только по числу позвонков, которое на Сахалине и Хоккайдо выше у северных популяций и у материковых популяций выше, чем у островных, расположенных на той же широте (табл. 51).
Миграции тайменя сходны с миграциями кунджи. Молодь, вышедшая из бугров, в июне — начале июля расселяется по прогреваемым мелководьям у песчаных и галечных кос после того, как с них произошла откочевка молоди других лососей. Начиная с годовалого возраста, молодь населяет в реках специфические стации: большие омуты или относительно глубокие и обширные плесы с медленным течением, с заиленным песчаным или галечным дном, с нависающими или упавшими в реку деревьями у подмытого берега. Молодь избегает участков с частым чередованием плесов и перекатов, поэтому ее нет в верховьях малых рек. В таких реках, как Танги и Первая Речка, для нагула молоди тайменя пригодны площади лишь в нижнем течении. В целом в малых реках площадь, используемая молодью тайменя для нагула, меньше нагульной площади молоди гольца, кунджи, симы или кижуча. По мере роста молодь тайменя расселяется по руслу реки, используя в Тыми и Поронае для нагула не только верхнее течение, но и равнинную часть основного русла. В период зимовок таймень скапливается на ямах отдельно от других видов рыб. В крупных реках (Тымь) молодь тайменя нагуливается до 5-7 лет, достигая длины 40-50 см, в малых реках (Танги, Первая Речка) молодь задерживается до 2-4 лет, вырастая до 10-25 см [Крыхтин и др., 1964; Гриценко и др., 1974; Гриценко, Чуриков, 19776]. О скате молоди тайменя из малых рек в сравнительно молодом возрасте свидетельствует также нахождение в озерах южного Сахалина нагульных особей в возрасте 1+ и 2+ [Завгородняя и др., 1964].
Скат молоди начинается предположительно после прохождения пика весеннего паводка и заканчивается во время летней межени. Протяженных морских миграций таймень не совершает, о чем свидетельствуют его неоднократные заходы в пресную воду в течение летнего нагула. В конце сентября — начале ноября таймень возвращается на зимовку в низовья рек или озера. В низовьях малых рек зимуют только мелкие особи (длиной до 25 см). Более крупные рыбы, видимо, заходят в низовья соседних крупных рек или в озёра. Так, например, известны места зимовки тайменя в озере Тунайча. До первого созревания таймень совершает зимовально-нагульные миграции река — море в течение 3-5 лет.
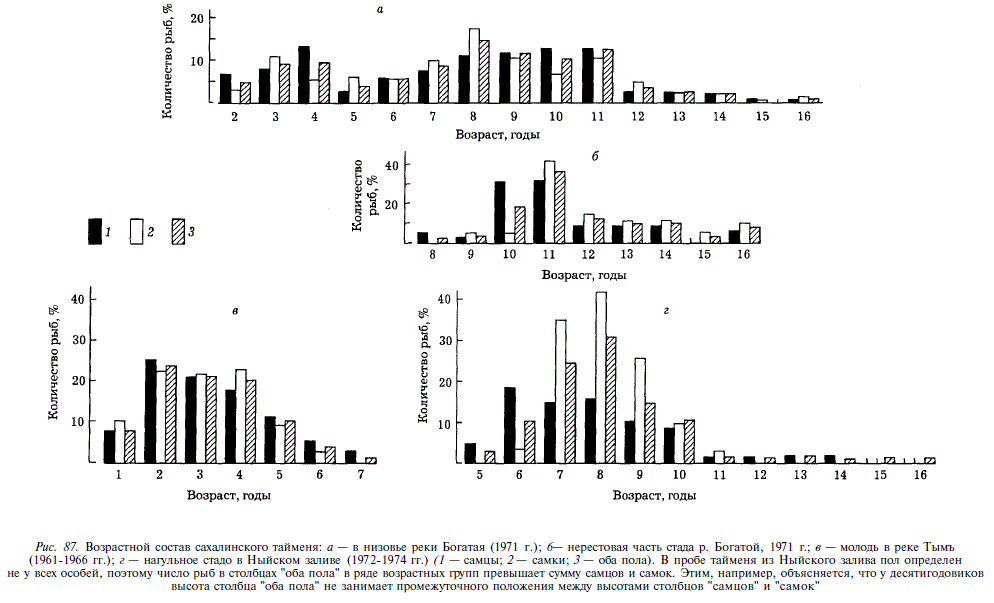
Нерест тайменя происходит весной и в начале лета в пик паводка и на спаде паводковой волны, в Японии в марте — апреле [Kimura, 1966], на Сахалине в мае — июне. Перед началом нерестовой миграции таймень, видимо, еще подо льдом на некоторое время скатывается в море. В крупных реках нерестилища расположены в верховьях, в малых реках — в нижнем и среднем течении. Нерест тайменя практически не изучен. Известно, что, подобно другим лососям, он закапывает икру в грунт, устраивая нерестовые бугры [Kimura, 1966]. Однако у впервые созревающих самок чешуя перед нерестом не погружается достаточно глубоко в кожу, что делает сомнительным возможность устройства ими гнезд в грунте. У повторно нерестующих особей преднерестовые изменения кожных покровов выражены сильно и вполне защищают рыбу от травм при копании грунта. После нереста таймень, как правило, сразу же скатывается из рек. Нерест у него неежегодный. Возраст тайменя определялся по чешуе. У рыб длиной более 110 см годовые приросты чешуи крайне незначительны и плохо различимы, в связи с чем возраст наиболее крупных рыб, по нашим определениям, видимо, существенно занижен. В Ныйском заливе встречались нагуливающиеся особи в возрасте 5-16 лет при численном преобладании 6-10-годовиков (рис. 87). Нерестовое стадо р. Богатая представлено 8-16-годовиками с преобладанием 10—11 -годовиков, в р. Набиль — 8-19-годовиками. Самцы тайменя начинают созревать на год раньше самок.
Рост тайменя в реке существенно отличается от роста в море (рис. 88). Начиная с 5 лет, у особей, нагуливающихся в Тыми, длина тела заметно меньше, чем у рыб, скатившихся в Ныйский залив. Отставание это увеличивается к 7 годам. Наиболее крупный из увиденных нами тайменей был добыт в Ныйском заливе: длина его 134 см, масса 37 кг, возраст более 16 лет.
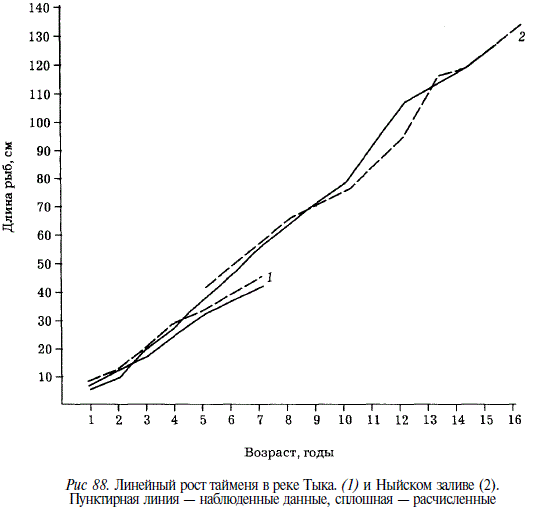
Рост тайменей, нагуливающихся в озерах и в море, существенно различается. Популяции, привязанные в период нагула к озерам, представлены более мелкими и тугорослыми особями, чем популяции, нагул которых происходит в море (река Богатая) и в больших заливах (например, в Ныйском) (табл. 52).
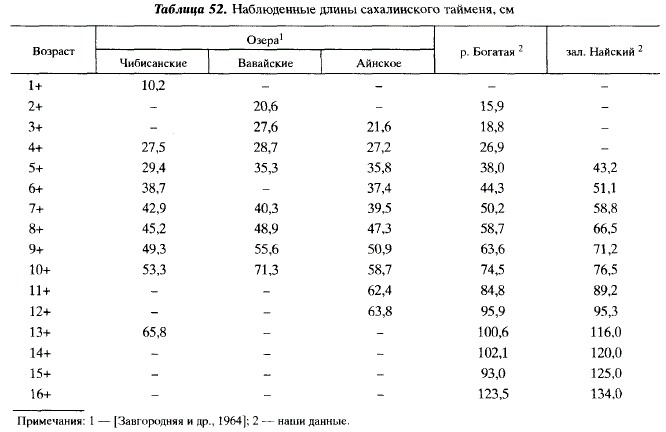
В Японии таймень созревает в более молодом возрасте и при меньших размерах, чем на Сахалине [Yamashiro, 1965; Kimura, 1966; Kawamura et al., 1983]. В настоящее время трудно сказать, чту это — результат влияния природных условий или антропогенной селекции в пользу рано созревающих рыб.
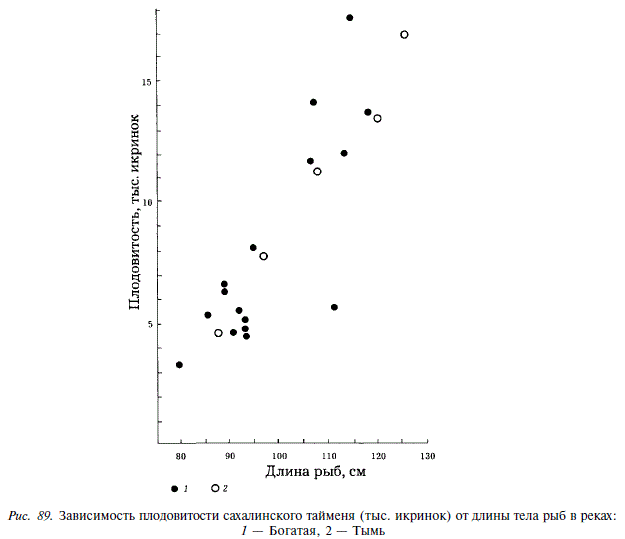
До наших работ сведений о плодовитости сахалинского тайменя в литературе не имелось. Мы определили плодовитость у 5 самок из реки Богатая, кроме того, в нашем распоряжении было 15 определений из реки Набиль. В первом случае она колебалась от 4670 до 16970 икринок, во втором случае — от 3380 до 17680 икринок, составляя в среднем 8300. Между длиной тела тайменя и плодовитостью существует положительная линейная зависимость (рис. 89). Диаметр икринок колеблется от 5,0 до 6,4 мм, средняя масса икринок различных особей — от 120 до 164 мг.
Представители рода тайменей по характеру питания являются хищниками. Однако, если сибирский таймень переходит на преимущественное потребление рыбной пищи с годовалового возраста, то сахалинский таймень питается преимущественно бентическими организмами [Крыхтин и др., 1964; Завгородняя и др., 1964; Ключарева, Световидова, 1968; Гриценко и др., 1974; Гриценко, Чуриков, 19776]. Эта закономерность не распространяется на сахалинского тайменя из рек Приморья, который, подобно сибирскому тайменю, начинает потреблять рыбу с первого года жизни по достижении длины тела 814 см [Бушуев, 1983]. Сеголетки сахалинского тайменя в период обитания на мелководьях питаются преимущественно бентосом. В рационе преобладают личинки хирономид и водяных клопов (табл. 53).
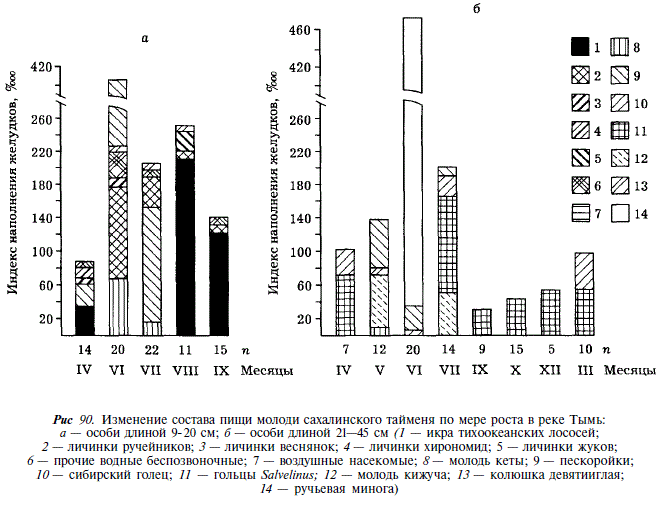
В Тыми молодь тайменя длиной 9-20 см потребляет личинок ручейников, веснянок, жуков, хирономид. Воздушных насекомых таймень, в отличие от других лососей, потребляет незначительно. Существенную часть рациона составляют мелкие пескоройки (длиной преимущественно до 10 см), икра тихоокеанских лососей и в отдельных случаях — молодь кеты и горбуши. Сезонные изменения наполненности желудков выражены слабо (рис. 90). Таймень длиной 21^45 см становится типичным хищником, потребляющим в течение круглого года преимущественно гольцов рода Salvelinus и сибирского гольца Nemachilus barbatulus toni. Наибольшие индексы наполнения желудка отмечаются в июне, когда таймень переходит на питание нерестующей ручьевой миногой. Из тихоокеанских лососей в желудках тайменей этой размерной группы встречается преимущественно молодь кижуча. Она потребляется тайменями в мае в старицах, где оба вида укрываются от паводка, и в основном русле среднего течения, где таймень потребляет нагульных пестряток, а также мигрирующих покатников (см. рис. 90).

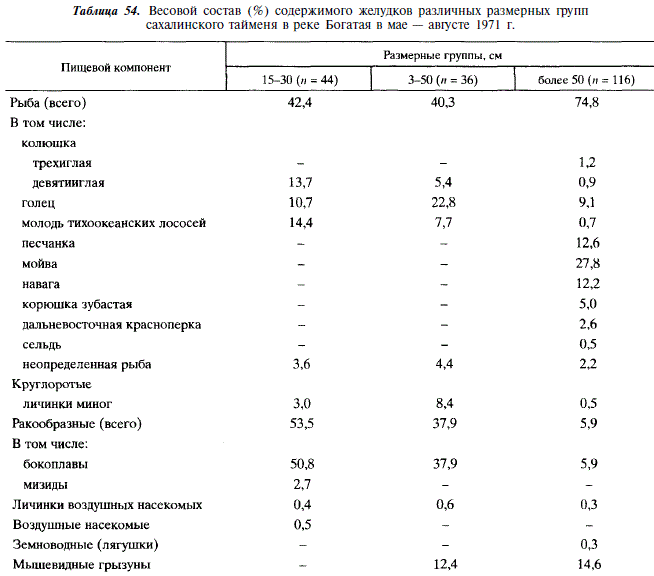
В низовьях реки Богатая молодь тайменя длиной до 50 см питается преимущественно рыбой (девятииглой колюшкой, гольцом, молодью тихоокеанских лососей) и ракообразными, среди которых преобладают бокоплавы. Существенную часть рациона составляют мышевидные грызуны (табл. 54).
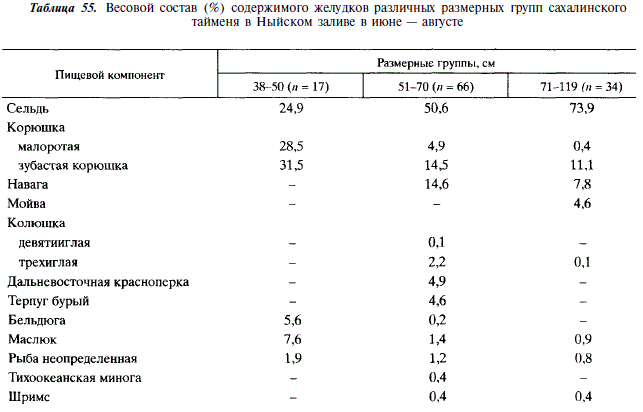
В период морского нагула таймень потребляет преимущественно рыбу в основном из толщи воды, однако в его рацион входят и придонные рыбы. Пищевой спектр при этом значительно расширяется (см. табл. 54, 55). Так, в Ныйском заливе в желудках тайменя встречается 12 видов рыб, в море, у реки Богатая, — 10 видов. В Ныйском заливе основу рациона составляют сельдь, малоротая и зубастая корюшки, а также навага. При этом по мере роста тайменя уменьшается доля потребления им корюшки и увеличивается — сельди. В период подходов к берегам нерестовой мойвы таймень переключается на питание ею. В море около устья Богатой таймень потребляет преимущественно мойву, песчанку, навагу, гольца и зубастую корюшку. Таким образом, основу рациона тайменя в период морского нагула составляют массовые пелагические и придонно-пелагические рыбы.
Сходный характер питания наблюдается у тайменя в озерах южного Сахалина и острова Кунашир [Завгородняя и др., 1964; Ключарева, Световидова, 1968]. У тайменя из озер острова Хоккайдо описано питание массовыми донными рыбами [Kawamura et al., 1983].
