Сима Oncorhynchus masu (Brevoort)
Сима обитает в северо-западной части Тихого океана, от Камчатки до южной оконечности Корейского полуострова [Берг, 1948; Machidori, Kato, 1984]. Наиболее численна она в бассейне Японского моря и сопредельных частях Охотского моря [залив Анива, юго-восточноный Сахалин, южные Курилы]. В пределах ареала этот вид представлен двумя сезонными формами: весенне-летней и летне-осенней, различающимися сроками нерестовой миграции, сроками и местами нереста [Иванков, Свирский, 1976; Семенченко и др., 1981а,б; Семенченко, 1984; Иванков и др., 1984а].
Согласно А.Ю.Семенченко [1985], каждая форма представлена комплексами локальных стад, которые характеризуются рядом особенностей биологии и населяют обширные регионы. Весенне-летняя форма населяет северную часть ареала, летне-осенняя — южную часть. Обе формы обитают совместно в ряде рек Приморья, например, в реке Киевка. Между сезонными расами были обнаружены различия по ряду морфометрических признаков [Семенченко, 1981а]. Однако дальнейшие исследования показали, что подобные различия по тем же признакам, существующие между локальными популяциями в пределах форм, часто превышают различия между формами [Иванков и др., 19846].
Высокая численность, симпатрическое обитание сезонных форм, большое разнообразие возрастных групп в нерестовом стаде, а также наличие самок, созревающих в пресной воде в Киевке, дают В.Н. Иванкову и соавторам [1984а] основание рассматривать эту реку в качестве центра внутривидовой дифференциации симы, а бассейн Японского моря — как зону экологического оптимума этого вида лососей. По нашему мнению, подобное предположение находит подтверждение в сводке С. Мачидори и Ф. Като [Machidori, Kato, 1984], где на рис. 5 наибольшее разнообразие в сроках нерестового хода наблюдается у симы, размножающейся в районе от 41 до 44° с.ш.
В Японии сима представлена проходной и жилой формами [Okada, 1960 и др.]. В реках России самки симы, созревающие в пресных водах, встречаются крайне редко и нерегулярно [Калмыкова, 1957; Крыхтин, 1962; Иванков и др., 1981], что, как мы считаем, свидетельствует об отсутствии репродуктивно обособленной жилой формы.
На Сахалине сима распространена повсеместно. По срокам нерестового хода и нереста ее следует отнести к весенне-летней форме. Нерестовый ход у нее начинается в период с конца апреля до начала июля, заканчивается с конца июля до конца августа (табл. 34). Однако в пределах весенне-летней формы мы выделяем у сахалинской симы две группы популяций, нерестящихся в сходные календарные сроки, но различающихся по времени захода в реки, степени зрелости половых продуктов в момент захода и, как следствие этого, по длительности периода дозревания в пресных водах. У производителей симы первой группы коэффициент зрелости у самцов обычно равен 0,7, у самок — 2,0; у производителей второй группы популяций — соответственно 2,0 и 4,5. Нерест обеих групп происходит с конца июля до начала сентября. Следовательно, созревание в реках у симы популяций первой группы длится около 3 мес. Следует отметить, что столь продолжительное пребывание производителей в реках до нереста уникально среди лососей рода Oncorhynchus и сходно с таковым у лососей рода Salmo. Эти группы популяций различаются также отношением к паводку в период нерестовой миграции. Первая заходит в реки в основном в период весеннего паводка, вторая — по его завершении. Первая обитает в реках западного Сахалина, залива Анива и юго-восточного Сахалина, от мыса Анива до мыса Терпения, вторая — в реках восточного Сахалина, впадающих в Охотское море к северу от мыса Терпения.

Основные нерестилища симы расположены по течению выше нерестилищ других тихоокеанских лососей. Большая часть производителей распределяется по капиллярам гидрографической сети острова, хотя имеются и нерестилища, расположенные в основном русле рек, где нерестовые площади симы перемежаются с нерестилищами горбуши [Канидьев, 1964; Смирнов, 1975]. Возможно, что ранний заход производителей в реки, приуроченный к весеннему паводку, является приспособлением, позволяющим симе попадать на те нерестилища, доступ к которым затруднен в условиях межени, особенно в маловодные годы.
Возрастная структура нерестовых стад симы,как и у кижуча, определяется соотношением особей, проведших в реке и в море разное количество лет. Однако у симы в нерестовом стаде присутствует еще один компонент — карликовые самцы, созревающие в пресных водах. В пределах ареала наиболее раннее созревание симы отмечается в возрасте 2j+, наиболее позднее — в возрасте 6j+. В реке молодь симы нагуливается до 3 лет, в море — до 4 лет [Крыхтин, 1962; Бирман, 1972; Иванков и др., 1984а; Семенченко, 1984]. Наибольшее разнообразие возрастных групп существует у симы южного Приморья. Подавляющее большинство особей симы повсеместно скатывается из рек на втором или третьем году жизни и нагуливается в море, подобно горбуше, около года. При этом в северных районах ареала наблюдается увеличение доли рыб старших возрастов, проведших в реках большее количество лет. Существующие представления о возрасте симы ставятся под сомнение В.В. Цыгиром [1970, 1988], который считает, что мнение о длительности морского нагула этого вида лососей объясняется неправильным прочтением регистрирующих структур. Имеющиеся наблюдения за ростом меченой симы в море и данные о возрасте меченых рыб, изложенные в ряде работ [Tanaka, 1965, 1970; Fukataki, 1970; Machidori, Kato, 1984; Mayama et al., 1985 и др.], дают основание считать, что сима проводит в море только одну зиму.
Отмеченное противоречие в определении возраста симы в малой мере касается сахалинских популяций. На Сахалине мигрирующие из моря производители симы представлены, как правило, особями в возрасте 2J+H 32+. Гораздо реже встречаются рыбы в возрасте 3,+, обнаруженные нами в реке Поронаи, и 42+, обнаруженные Ф.Н. Рухловым и соавторами [1976] в реке Набиль (табл. 35). Отмеченная выше тенденция увеличения доли рыб старших возрастов в северных популяциях проявляется и в пределах Сахалина. Это увеличение происходит за счет того, что в районах с более холодным климатом и сравнительно коротким периодом вегетации скат молоди происходит в более старшем возрасте. Карликовые самцы симы на Сахалине обычно созревают в возрасте 1+ и 2+, изредка встречаются особи в возрасте 3+. Среди них доля рыб младшего возраста, как и среди проходных самцов, выше в южной части острова и на япономорском побережье, чем в северной части и на охотоморском побережье (табл. 36). Созревания в возрасте 0+, обычного в Японии [Machidori, Kato, 1984], на Сахалине не отмечается. У симы карликовые самцы не погибают сразу же после нереста подобно другим тихоокеанским лососям. В течение сентября — октября они питаются, а гонады у них переходят во II стадию зрелости [Крыхтин, 1962; Воловик, 1963; Иванков и др., 1977; наши данные]. В.Н. Иванков с соавторами [1977] высказывают предположение, что карликовые самцы могут нерестовать неоднократно.
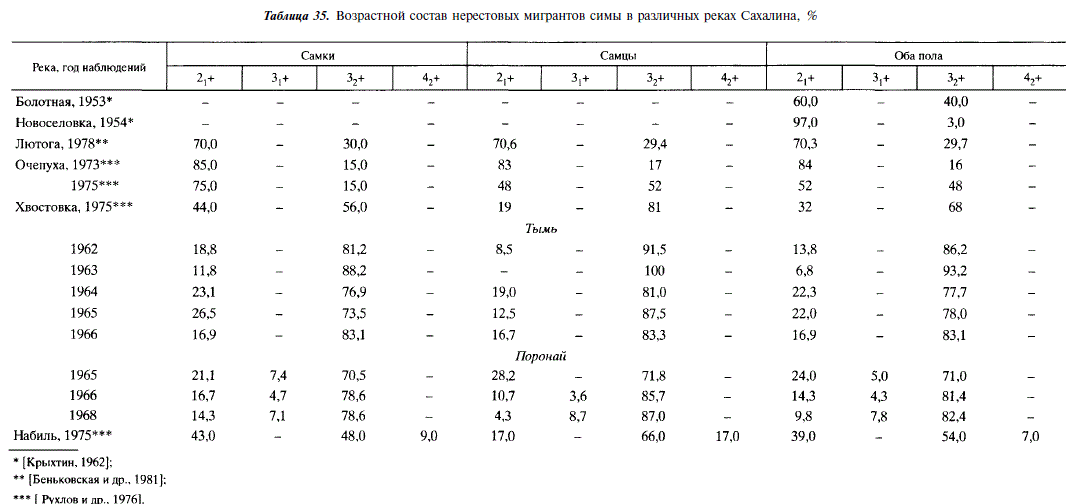

Среди нерестовых мигрантов симы повсеместно преобладают самки. Согласно С. Мачидори и Ф. Като [1984], проходная часть некоторых популяций острова Хонсю практически на 100 % представлена самками. По данным этих авторов, в Японии доля проходных самцов минимальна в районах с теплым климатом. Так, на острове Хоккайдо доля проходных самцов значительно уменьшается в районах, обогреваемых теплым Цусимским течением (табл. 37).

В сахалинских популяциях симы среди производителей-мигрантов также преобладают самки, хотя в отдельные годы соотношение полов бывает равным 1:1. Тенденция увеличения доли самок в южных районах острова прослеживается, если сравнивать данные только по тем рекам, где собран наиболее представительный материал (Таранай, Лютога, Поронай, Тымь). У молоди, нагуливающейся в реках, за редким исключением, соотношение самцов и самок примерно 1:1 (табл. 38).
Среди покатной молоди, как и среди мигрантов производителей, преобладают самки. В Тыми в 1965 г. их доля составила 63 %, в Богатой в 1971 г. — 55 %. На нерестилищах симы численно преобладают карликовые самцы. Обычно около самки находятся один проходной самец и несколько (как правило, не больше 7-9) карликовых. На нерестилищах в верховьях ручьев в нересте часто участвуют только карликовые самцы.
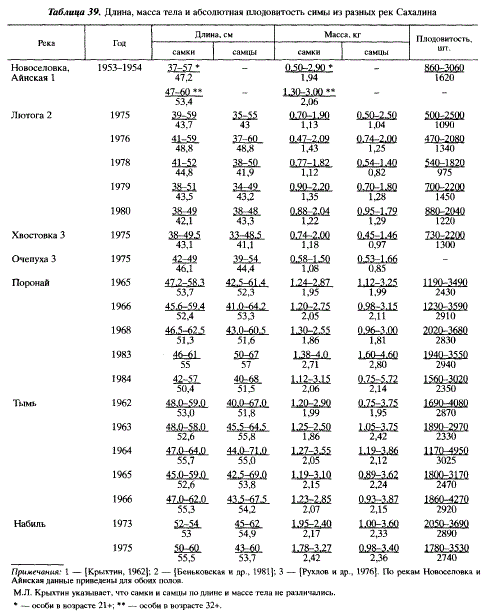
Размеры и плодовитость мигрирующих производителей симы в одной реке значительно колеблются по годам (табл. 39). Однако эти колебания находятся в пределах, характерных для отдельных районов. Мы выделяем три таких района. Так, сопоставляя наши данные с данными, приводимыми в сводке С. Мачидори и Ф. Като [Machidori, Kato, 1984], можно отметить большое сходство в размерах тела между японской симой, симой залива Анива и юго-восточного побережья Сахалина. В этих районах средняя длина производителей симы обычно не превышает 50 см, масса — 1,5 кг, плодовитость — 1500 икринок. Другую однородную группировку составляет сима Пороная и рек северо-восточного Сахалина, у которой средняя длина бывает не менее 50 см, а масса — не менее 1,8 кг (чаще всего — 52-54 см и 2,0-2,2 кг соответственно), средняя плодовитость составляет 2300-2900 икринок. Сима западного Сахалина (реки Айнская, Новоселовка) занимает по этим показателям промежуточное положение между двумя упомянутыми выше группировками, по средней длине и массе тела она ближе к симе северо-восточного Сахалина, по плодовитости — к симе залива Анива и юго-восточного Сахалина. С симой из двух последних районов западносахалинскую симу сближает также наличие мелких особей длиной до 40 см, массой менее 700 г, плодовитостью менее 1 тыс. икринок, которые отсутствуют на северо-востоке острова. Как известно, наиболее крупная плодовитая сима обитает в Приморье, где, в свою очередь, доминирует по размерам и плодовитости осенняя форма [Воробьев, 1926; Семенченко, 1984; Иванков и др., 1984а]. Сима северо-восточного Сахалина приближается по размерам и плодовитости к весенне-летней форме приморской симы и сходна с амурской симой [Двинин, 1956].

Молодь симы выходит из грунта в марте — апреле. В отличие от кижуча расселение ее в первый год жизни идет только вниз по течению. Расселение это довольно ограниченно. Так, в ключе Холодный (бассейн Пороная) вблизи нерестилищ симы расселяющиеся сеголетки в большом количестве попадаются в ловушку при учете молоди горбуши и кеты. В верхнем течении Тыми сеголетки попадаются в ловушки единично, а в среднем течении не попадаются вообще.
Сезонные миграции молоди симы происходят по следующей схеме. Мальки с апреля по июнь держатся на мелководьях у берегов, осередков и в закосьях. В июле они переходят на более глубокие места, заселяя перекаты и плесы. В октябре молодь мигрирует вниз по течению, где концентрируется на зимовальных ямах в низовье нерестовых притоков или в основном русле, а также заходит в лимнои реокрены придаточной системы, не замерзающие в течение зимы. В малых реках (Первая Речка, Ударница, Айруп) молодь мигрирует на зимовку на плесы и омуты нижнего течения. Перезимовавшая молодь частично до весеннего паводка, частично после него поднимается вверх по течению в район нерестилищ, откуда либо скатывается в море в возрасте 1+, либо повторяет сезонный цикл миграций, скатываясь после второй зимовки. Предпокатная молодь после зимовки не поднимается по течению так далеко, как пестрятки.
Наиболее крупные особи как во время летнего нагула, так и во время зимовки населяют наиболее крупные водотоки в пределах бассейна реки, а в пределах самих водотоков — наиболее глубокие и обширные перекаты. Это хорошо видно из материалов М.Л. Беньковской и соавторов [1981], в бассейне Лютоги собравших в один из дней сентября пробы молоди из трех различных по размерам водотоков. Длина и масса молоди симы в этих водотоках, расположенных в порядке убывания их размеров, были соответственно 8,40 см и 7,14 г; 7,90 см и 6,21 г; 6,80 см и 3,82 г. По данным А.Ю. Семенченко [19816], у молоди симы в пределах возрастных классов образуются размерные группы, в основе которых лежат наследственные особенности роста, а также доминантно-иерархические отношения в местах нагула.
В бассейне Тыми, где имеются обширные незамерзающие участки, рост молоди симы происходит круглогодично. Однако наиболее интенсивным он бывает с июня по сентябрь. С конца сентября рост замедляется. Очередное его ускорение у 2-леток начинается в июне, а у 3-леток, готовившихся к скату, — на полтора месяца раньше — в апреле (рис. 56). Скат симы подробно изучен в Японии. Нами в 1971 г. было проведено первое в отечественных водах специальное изучение ската молоди этого вида в р. Богатая, которое до сегодняшнего дня остается одним из немногих. Для японской симы экспериментально установлено, что длина тела 6-7 см является пороговой для начала смолтификации. Однако большинство смолтов-сеголеток вновь превращаются в пестряток и скатываются лишь на следующий год [Kurokawa et al., 1985]. Возраст смолтификации во многом определяется условиями нагула молоди. Так, при высокой ее плотности в реках увеличивается количество мигрантов в возрасте 2+ [Sugiwaka, Kajima, 1979]. Новолуние и выпадение дождей в определенные сезоны года являются важнейшими сигнальными факторами среды, вызывающими покатное поведение у молоди симы [Izumi et al., 1984]. Это хорошо согласуется с тем, что концентрация тиросина — гормона, участвующего в осуществлении осморегуляции, — бывает наибольшей в мае, в периоды новолуния [Yamauchi et al., 1984].
Скат симы в Японии происходит с конца апреля по начало июня. В южных районах он начинается раньше, чем в северных [Sano, Abe, 1967; Sugiwaka, Kajima, 1979; Kubo, 1980; Machidori, Kato, 1984]. В бассейне Японского моря наряду с весенне-летним отмечается незначительный по численности осенний скат симы [Крыхтин, 1962; Иванков и др., 1984а].
На Сахалине экология ската симы имеет ряд региональных особенностей. В южных районах острова молодь скатывается с конца мая до I декады июня , в северных районах — с I декады июля по I декаду августа (табл. 40, см. рис. 51). Соответственно в южных районах скат начинается в период прохождения пика весеннего паводка, а основная масса молоди скатывается на спаде паводковой волны (реки Лютога, Очепуха). В реках северовостока (Тымь, Богатая) сима скатывается после весеннего паводка при расходах воды, приближающихся к меженным. Соответственно в первых двух реках скат симы совпадает по времени со скатом молоди горбуши, во вторых двух — происходит после его окончания.

Подобное смещение сроков ската обеспечивает для молоди симы разных рек попадание в сравнительно сходные температурные условия в море. Так, если поверхность воды в заливах Анива и Мордвинова прогревается в июне до 7-10 °С (обычно до 8-9 °С), то у северо-восточного Сахалина такой прогрев наступает лишь в июле. В пределах острова сима скатывается в сходных температурных условиях в реках. Скат начинается при прогреве воды до 710 °С, основная масса скатывается при 10-12 °С. Однако в среднем и нижнем течении крупных рек (Тымь, Лютога) температура вода во время ската достигает 18-20 °С.
Более 90 % особей симы на Сахалине скатывается в возрасте 1+ и 2+, при этом доля старших рыб увеличивается на севере острова. Сеголетки составляют незначительную часть мигрантов. Они были обнаружены лишь в Найбе [Воловик, 1963] (табл. 41).
Размеры покатной молоди симы обычно имеют незначительные различия. В большинстве рек средняя длина покатников колеблется от 11 до 14 см. Особенности размерного состава проявляются в том, что особи старших возрастов значительно крупнее младших. Так, в реках Лютога и Найба масса 3-леток в 1,5 и 2 раза превышала массу 2-леток (см. табл. 41).

Покатники симы в лимане Богатой и в устье Тыми практически не встречались в желудках потенциальных хищников (сахалинского тайменя, кунджи, гольца). Слабое воздействие хищников на покатную молодь симы отмечалось также в северном Приморье [Горяинов, 1985].
Заключая рассмотрение ската молоди симы, следует отметить, что по особенностям ее экологии в пределах Сахалина выделяются те же две группы популяций, что и по особенностям нерестовой миграции: 1 — популяции западного побережья, залива Анива и юго-восточного побережья; 2 — популяции северо-восточного побережья.
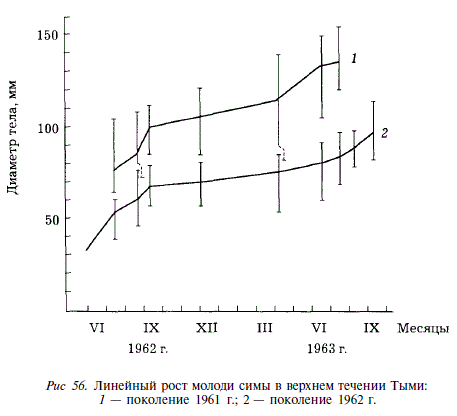
Предыдущими исследованиями было установлено, что молодь симы обладает весьма широким спектром питания. Так, СП. Воловик [1964] обнаружил в ее желудках более 150 различных пищевых компонентов; при этом определение большинства из них проводилось до рода или более крупных таксономических единиц (до семейств или класса). Сезонные особенности питания симы исследовались нами в бассейне Тыми. Наибольшие индексы наполнения желудков у всех возрастных групп отмечаются с июня по сентябрь. Основу рациона составляют в это время две группы пищевых организмов: бентические личинки насекомых (хирономид, веснянок, поденок, ручейников) и имаго воздушных насекомых, упавшие в воду. Воздушные насекомые встречаются в желудках молоди симы до начала октября, после чего вплоть до июня молодь питается бентосом. Существенным компонентом рациона является икра кеты и горбуши, которую наиболее крупные сеголетки начинают потреблять в августе. Этот корм потребляется молодью с осени до начала лета (рис. 57). Перечисленные группы личинок насекомых потребляются молодью симы на протяжении всего пребывания в реках, однако по мере роста молодь поедает все более крупные формы. Эта особенность питания симы отмечалась М.Л. Крыхтиным [1962]. В рационе 2-леток, по сравнению с таковым сеголеток, возрастает доля личинок бабочек и жуков. Покатная молодь кеты и горбуши единично встречается в желудках симы в течение июня и июля. Среднее количество экземпляров в одном желудке по разным пробам не превышает 0,2.
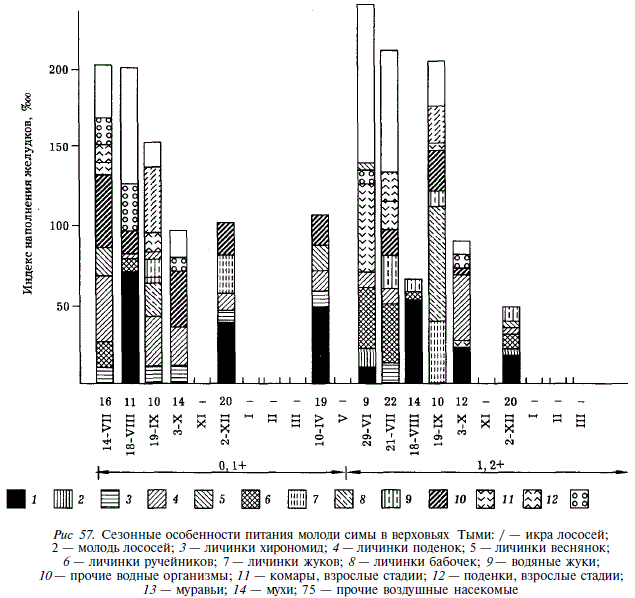
Для изучения биотопических особенностей питания симы использовались пробы, собранные в июле — начале августа. Повсеместно симе свойственно сочетание бентофагии и потребления воздушных насекомых. Однако ведущие компоненты бентоса существенно различаются как в различных реках, так и в стациях в пределах одной реки (рис. 58). Так, в Первой Речке молодь потребляет значительное количество личинок хирономид и стрекоз, что в малой степени свойственно ей в реках восточного Сахалина. В маленьких реках юго-восточного Сахалина Айруп и Ударнице ведущими компонентами рациона являются ракообразные (бокоплавы и ветвистоусые). В бассейне Богатой среди потребляемых организмов преобладают личинки веснянок, поденок и ручейников, а в лимане этой реки начинают доминировать ракообразные, 95 % массы которых составляют бокоплавы и 5 % — кумовые раки. В бассейне Тыми наряду с личинками веснянок, поденок и ручейников важную роль в питании молоди симы играют личинки бабочек и жуков. В старице Тыми, где нам удалось добыть несколько экземпляров двухлеток симы, они питались преимущественно водяными жуками. Таким образом, биотопические особенности питания молоди симы определяются особенностями фауны беспозвоночных в населяемых ею водотоках.

Суточная ритмика пищевой активности молоди симы изменяется по мере роста рыб. Мальки, обитающие после выхода из грунта на мелководьях, потребляют корм круглосуточно, имеют постоянно высокую наполненность желудков, на фоне которой отмечаются три подъема: с утренних часов до полудня; послеполуденный подъем с14до18чи подъем в вечерних сумерках, с 20 до 22 ч. Минимальное наполнение желудков приходится на ночные часы (рис. 59) [Воловик, 1961]. У сегелеток-пестряток наполнение желудков ниже, чем у мальков, а перепады наполнения в пределах суток выражены сильнее. Отмечаются два периода повышенной пищевой активности в утренние (6 до 10) и вечерние (с 18 до 00) часы (см. рис. 59,6) [Воловик, 1961]. У пестряток 2-леток и серебрянок 2и 3-леток мы наблюдали дальнейшее уменьшение среднего за сутки наполнения желудков и увеличение амплитуды содержания пищи в период подъема и спада активности питания. Как и в предыдущем случае, достаточно хорошо выражены два подъема пищевой активности: первый — в утренние часы (с 5 до 9), второй — в дневные и вечерние (с 11 до 23). Период замедления потребления пищи в данном случае короче (см. рис. 59,6). В лимане реки Богатая накануне выхода в море пищевая активность покатников бывает наибольшей в вечерние и ночные часы за счет потребления ракообразных, поднимающихся в толщу воды. Утренний подъем пищевой активности, отмечаемый в предыдущих случаях, отсутствует, однако наблюдается незначительный подъем в дневное время с максимумом в 14 ч, за счет потребления воздушных насекомых, поедаемых молодью с поверхности воды (см. рис. 59,г).
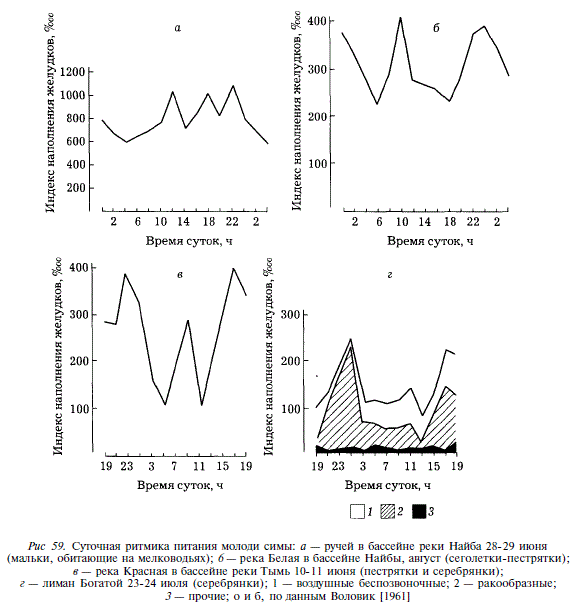
Таким образом, на протяжении пребывания молоди симы в пресных водах отмечается уменьшение среднего за сутки наполнения желудков и увеличение его колебаний в течение суток. В значительной мере суточная активность симы определяется наличием и доступностью корма. В целом же ей наиболее свойственно питаться в часы утренних и вечерних сумерек.
Сопоставляя особенности питания молоди симы и кижуча, нельзя не заметить их большого сходства в местах совместного обитания. Однако ряд особенностей биологии этих видов ослабляет их взаимодействие на почве потребления сходного корма. СП. Воловик [1964] обнаружил, что сеголетки симы и кижуча в разное время обитают на мелководьях. Наши исследования показали, что 2и 3-летки этих видов имеют специфическое распределение по различным стациям.
Сима по дальневосточным меркам считается второстепенным объектом промысла. Добыча ее была наиболее развита в северном Приморье, где в низовьях Амура, на югозападном Сахалине и в заливе Анива в конце 30-х годов вылавливалось от 0,6 до 1,2 тыс. т. На юго-западном Сахалине вылов симы в отдельные годы составлял около 40 % от общей с горбушей добычи [Двинин, 1952]. В последние 20 лет отечественный вылов симы не превышает 1 тыс. т, при этом в большинстве случаев рыбаки сдают ее совместно с горбушей, не выделяя в статистике вылова. Основная масса симы добывается Японией. Так, в 70-е годы ее вылов колебался от 3,0 до 4,2 тыс. т. Из этого объема лишь 1,5-2,5 % приходилось на береговой промысел, основное же ее количество добывалось в море во время зимовки [Machidori, Kato, 1984]. Поскольку в Японском море и прилежащих к Японии водах Тихого океана зимуют практически все популяции симы [Двинин, 1957; Бирман, 1972; Никифоров, 1985; Tanaka, 1965; Machidori, Kato, 1984], естественно, что столь высокие уловы обеспечиваются в первую очередь воспроизводством этого вида лососей в водах России, где находятся основные нерестилища. Рассчитывать в этих условиях на увеличение российского вылова едва ли возможно. Факторами, ограничивающими воспроизводство симы в отечественных водах, являются браконьерский вылов производителей и любительский лов молоди. Численность симы в нерестовых реках трудно поддается определению. На основании нерегулярно проводимых визуальных учетов можно считать, что в крупные реки Сахалина (Тымь, Поронай) заходит не более 20 тыс. особей, в средние — порядка 1-2 тыс. Наиболее высокой была численность в Лютоге, где А.Н. Канидьевым [1968] было учтено в 1963 г. 180 тыс. производителей. Нам представляется, что увеличение численности симы возможно лишь при организации интенсивного искусственного разведения, которое должно сочетаться с введением заповедного режима на ряде рек и установлением ограничений на японский морской промысел. Учитывая малую численность и большую дисперсность стад симы, можно рекомендовать в качестве оптимального способа ее использования лицензионный спортивный лов.
