Тихоокеанская сельдь Clupea pallasi Valenciennes
По современным представлениям, существуют две экологические формы тихоокеанской сельди — морская и лагунная (озерная), каждая из которых включает ряд локальных стад, являющихся единицами хозяйственного использования и научного исследования [Фролов, 1949,1950,1954,1968; Веденский, 1950; Пробатов,
Фролов, 1951,1958; Румянцев и др., 1958; Рыбникова, 1985]. К морской форме относят сельдь залива Петра Великого, сахалино-хоккайдскую, охотскую, гижигинско-камчатскую, пенжинскую, корфокарагинскую; к лагунной форме — сельдь сахалинских озер Тунайча, Айнского, заливов северо-восточного Сахалина и Сахалинского залива, хоккайдских озер Ноторо, Аккеш, Хинума, камчатских озер Нерпичье, Култучное, Топатское. К лагунной форме относят также декастринскую сельдь и сельдей заливов Анива и Терпения. Между морскими и лагунными сельдями существуют значительные различия в образе жизни. Лагунные сельди по сравнению с морскими нерестятся в более поздние календарные сроки при относительно низкой солености и высокой температуре воды, не совершают протяженных и продолжительных миграций, заходят на зимовку в опресненные водоемы, созревают в сравнительно молодом возрасте и при малых размерах тела. Предельная длина тела, возраст и плодовитость у них относительно невелики (табл. 6).
Далеко не все стада тихоокеанской сельди изучены достаточно подробно. Дефицит информации подчас делает затруднительным отнесение того или иного стада сельди к одной из двух упомянутых форм. В связи с этим на сегодняшний день, возможно, будет уместнее говорить о существовании некоего ряда экологически различающихся популяций, на одном конце которого находятся типично морские сельди, на другом — типично лагунные. В наибольшей мере черты морской формы свойственны сельди залива Петра Великого, черты лагунной формы — стаду озера Ноторо (Хоккайдо), где имеются две группы особей: мигранты и оседлые, не покидающие его пределов [Каппо, 1981]. Среди сахалинских сельдей наиболее выраженными свойствами лагунной формы обладают популяции озер Тунайча и Айнское. Эти сельди нерестятся в солоноватой озерной воде (4 — 7 %о), молодь их в первый год жизни не покидает озер. В дальнейшем рыбы в течение летних месяцев нагуливаются в море, в районе озер, а осенью возвращаются в них на зимовку [Пробатов, Фролов, 1951; Румянцев и др., 1958; Фролов, 1968].
Сельди декастринская, анивская и залива Терпения занимают промежуточное положение между морскими и лагунными. Они нерестятся в условиях меньшей солености, чем морские, но большей, чем типично озерные. Места зимовки стад точно не установлены, но, исходя из общей гидрологической обстановки в районах их обитания, соленость в местах зимовки должна быть выше, чем у типично озерных сельдей. В этой связи интересно отметить, что декастринская и анивская сельди осенью на 15 — 40 сут. подходят к берегам в воды с пониженной соленостью, прекращая в это время питаться, после чего откочевывают на зимовку [Пробатов, Козлов, 1954; Дружинин, 1957].
Нами изучалась сельдь Ныйского залива. Предыдущими исследователями было установлено, что она нерестится в заливе и в течение всего лета нагуливается в море неподалеку от залива, заходя в него неоднократно на протяжении летнего нагула. Зимует сельдь в заливе и прилегающих участках моря, опресняемых стоком Тыми [Амброз, 1931а, б; Веденский, 1950; Фролов, 1968].
Существует другое мнение относительно миграций сельди северо-восточного Сахалина. З.Г. Иванкова и Б.М. Козлов [1968] на основании данных мечения и анализа ряда фенотипических признаков пришли к выводу, что после нереста сельдь этого стада мигрирует на юг в залив Терпения, где нагуливается до осени, после чего возвращается на зимовку в заливы северо-восточного Сахалина. Сельдь, меченную в Ныйском заливе, вторично вылавливали также у берегов Камчатки и в заливе Петра Великого [Андреев, 1968]. Нам эти точки зрения не кажутся взаимоисключающими, ибо, по аналогии с сельдью озера Ноторо, в данном случае также возможно существование оседлых особей и мигрантов. Это предположение подтверждается нахождением в летние месяцы молоди сельди в желудках сахалинского тайменя, добытого в устье реки Богатая, расположенном примерно посередине между заливами Ныйский и Терпения. Миграции сельди должно способствовать существование вдоль побережья зоны, опресняемой речным стоком.
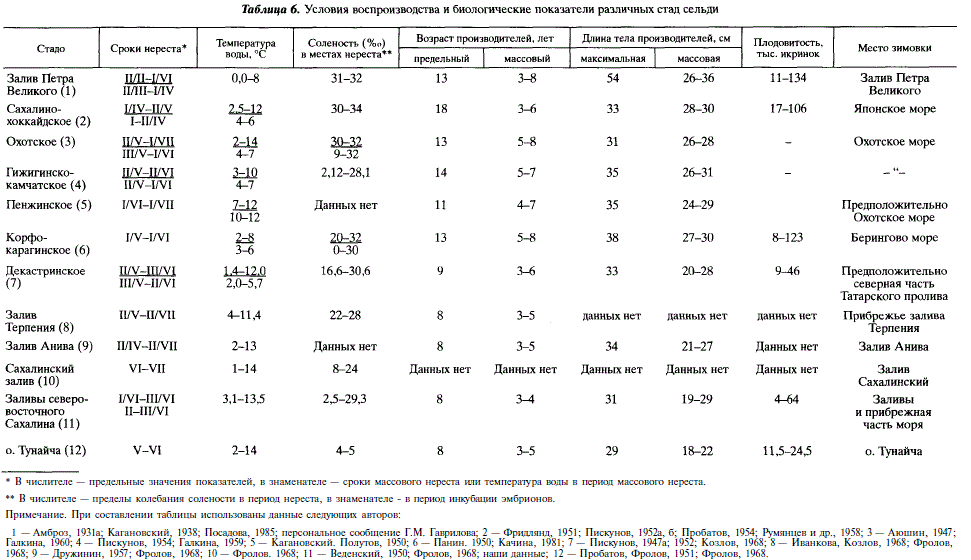
Об условиях нереста тихоокеанских лагунных сельдей имеются лишь отдельные сведения общего характера [Фролов, 1949,1950,1968]. Летом 1972 и 1973 гг. мы попытались восполнить этот пробел. Для наблюдения за подходами и изменением биологических показателей рыб проводились регулярные обловы стандартными жаберными сетями. В 1972 г. контрольные сети были поставлены 24 мая. До 17 июня суточные уловы не превышали 10 экз. С 18 по 23 июня отмечался массовый заход сельди на нерест. В дальнейшем отмечалось еще два массовых захода — 5 и 16-18 июля (рис. 5). Сопоставление величины уловов контрольных сетей с колебаниями уровня воды в заливе показало, что массовые заходы в 1972 г. отмечались в периоды квадратурных приливов (см. рис. 5). В это время суточные характеристики гидрологического режима относительно постоянны. Соленость, уровень и температура воды колеблются в гораздо меньших пределах, чем в период сизигий. Так, в середине июня в период квадратур суточный перепад солености составил 2,6 %о (14,4—17,0), температуры воды — 0,8 °С (7,6-8,4). В конце июня, в период сизигий, эти показатели составляли соответственно 14,4 %о (3,9-18,3), 4,6 °С (6,4-11,0) и 1,3 м.
Привязанность нереста к периодам квадратур отмечается и у атлантической сельди Clupea harengus (L), нерестующей в мелководных заливах [Clarke, King, 1985]. Это, видимо, связано с тем, что во время оплодотворения и начала эмбриогенеза стабильные условия среды наиболее благоприятны для сельди. В отличие от вышеописанного, нерест корфо-карагинской сельди происходит преимущественно в период сизигий [Панин, 1946; Качина, 1981]. Объясняется это тем, что резкие перепады уровня способствуют освобождению нерестилищ ото льда и прогреву воды. Вероятно, для тихоокеанских морских сельдей фотопериод и температура воды в большей мере определяют начало нереста, чем колебания солености и уровня воды.
На основании поимки производителей с текучими половыми продуктами было установлено, что нерест сельди в Ныйском заливе в 1972 г. происходил с 9 июня по 26 июля. Интенсивность его на протяжении этого времени не была одинаковой вследствие колебаний числа заходов и различной доли в них зрелых рыб. До 18 июня 80 % всех пойманных рыб имели половые продукты в IV и V стадиях зрелости. Во время первого, наиболее массового, захода таких рыб было 78 %, во втором и третьем заходах — соответственно 14 и 7 %. Сопоставляя относительную численность заходов и долю в них особей с гонадами в IV и V стадиях зрелости, можно предположить, что в период с 18 по 25 июня отнерестилось 70-80 % нерестового стада сельди.
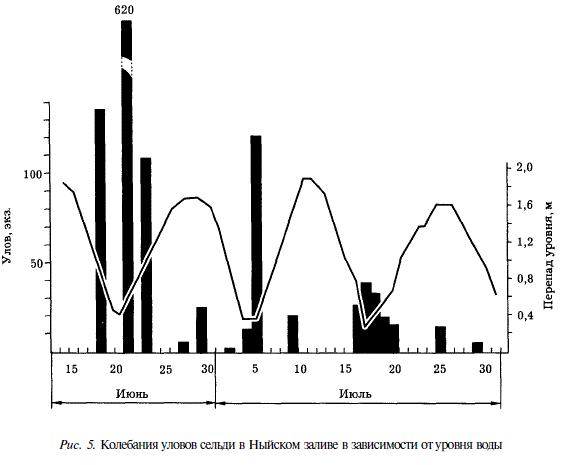
От начала к концу нереста наблюдалось закономерное изменение биологических показателей производителей. Сначала в нерестовом стаде значительная доля приходилась на старшие возрастные группы. Так, в уловах с 25 мая по 17 июня было довольно много 6-7-годовиков, а в последнем подходе самой старшей возрастной группой явились 5-годовики (рис. 6).
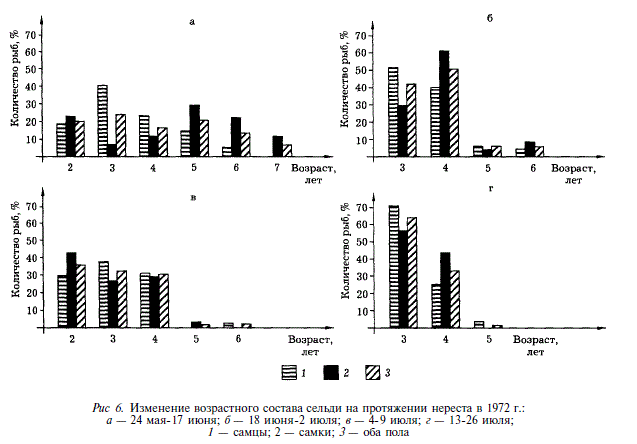
Соотношение полов от начала к концу нереста также изменилось: если в его начале самки составляли 35-39 % общей численности, то в конце — 45-47 %. В целом для сельди Ныйского залива характерно относительное преобладание числа самцов на всем протяжении нереста.
Длина нерестовой сельди в 1972 г. колебалась от 18 до 30 см с преобладанием рыб длиной 19-23 см. Средняя плодовитость равнялась 29,3 тыс. икринок при колебаниях от 3,8 до 64,0 тыс. По сравнению с сахалино-хоккайдской сельдь Ныйского залива менее плодовита: для первой указывается средняя плодовитость 53,7 тыс. икринок [Фридлянд, 1951].
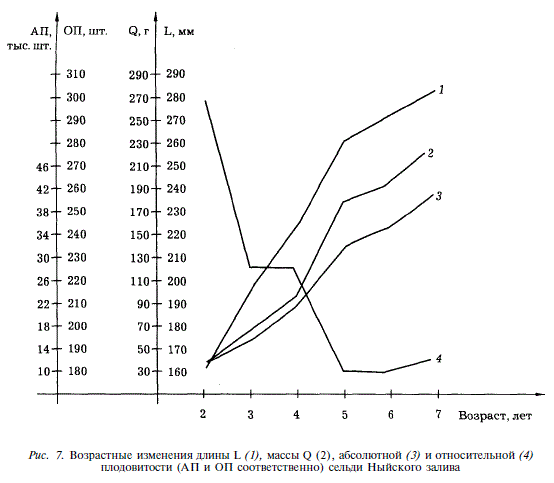
За период от 2 до 7 лет происходит примерно четырехкратное увеличение абсолютной плодовитости и семикратное увеличение массы (рис. 7). С этим связано снижение относительной плодовитости рыб с возрастом.
В 1973 г. первый массовый заход сельди отмечен 17 июня. В дальнейшем наблюдалось еще два захода — с 22 по 26 июня и с 6 по 9 июля. В отличие от предыдущего года, только второй и третий заходы пришлись на период квадратур, первый заход был в период сизигий. Обследование нерестилищ показало, что основным нерестовым субстратом сельди является зостера, как это свойственно малопозвонковым сельдям в целом. Икра была отложена на листьях зостеры в один слой. Среднее количество икринок на одном листе длиной 30-40 см составило 64 шт. при колебаниях от 0 до 231 (просматривалось по 20 листьев). В дальнейшем по мере развития эмбрионов среднее количество икры в кладках уменьшалось: через 3 дня оно снизилось до 49 шт. при колебаниях от 0 до 184, через 7 дней — до 14 икринок (0-89).
22 июня была взята проба икры для определения стадий развития. Из 55 икринок, составлявших пробу, развивающихся было 81 %, неоплодотворенных — 4 %, пустых оболочек — 15 %. Из развивающихся икринок 2 % находились на стадии образования бластодиска, 2 % — на стадии морулы, у остальных 96 % происходило дробление бластодиска (насчитывалось от 8 до 32 бластомеров). Вторая проба была взята на том же нерестилище 1 июля. Из 52 икринок 86 % составляли развивающиеся, 10 % — неоплодотворенные и 4 % пустые. При этом 53 % развивающихся эмбрионов находилось на стадии позднего эмбриогенеза, 47 % — накануне выклева. Через несколько дней, 4 июля, икру на нерестилищах обнаружить не удалось. Однако на одном из островов залива была найдена выброшенная прибоем зостера с выметанной на ней икрой. Из 235 икринок развивающиеся составляли 68 %, неошюдотворенные — 29 % и пустые — 3 %. Вся развивающаяся икра находилась на стадии мелкоклеточной морулы и, следовательно, была выметана производителями более поздних подходов. Исходя из этих наблюдений, можно считать, что развитие эмбрионов сельди с момента нереста до вылупления длится 10-12 дней.
За период развития икры температура воды в заливе колебалась от 3,1 до 13,5° С с максимальным внутрисуточным перепадом 6,4°С (5,4-11,8). Соленость изменялась от 2,5 до 29,3 %с с максимальным внутрисуточным перепадом 21,0 %о (2,5-23,5). При минимальном уровне воды в заливе глубина на нерестилищах составляла 0,2-0,3 м, при максимальном — 2,2-2,3 м.
Во время отливов листья зостеры до половины своей длины оказываются на поверхности воды. Из-за ветров, дующих почти постоянно, и волнения они находятся в движении, что приводит к отрыву икринок от субстрата. Это является одной из причин отмеченного выше уменьшения среднего количества икры в кладках на протяжении эмбрионального периода.
Поскольку в естественных условиях обитания сельди сильное изменение параметров среды, в частности солености, является нормой, представляло интерес выяснить особенности развития икры при различных константных значениях солености. С этой целью была поставлена серия опытов по инкубации икры в воде соленостью 2,6; 15,0; 25,0 и 29,0 %с. Опыт был начат 21 июня и закончился 10 июля, по прошествии суток после вылупления последнего эмбриона.
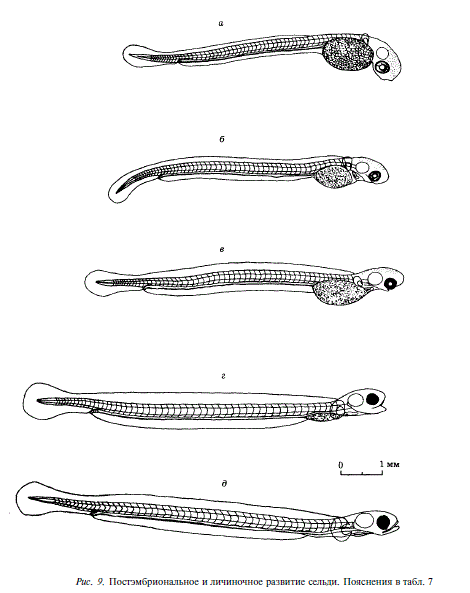
Вылупление личинок наблюдалось во всем диапазоне солености. На фиксированном материале было прослежено развитие сельди при 2,6 и 29,0 %0 и сделаны рисунки эмбрионов, развивавшихся при 2,6 %о (рис. 8, 9, табл. 7). На этом уровне солености мы остановились потому, что он необычен для развития икры сельдей рода Clupea. Картина развития при 2,6 и 29,0 %о была сходной. В опыте с соленостью 29,0 %0 развитие эмбрионов шло с запозданием примерно на сутки по сравнению с изображенным на рис. 8, 9 и указанным в табл. 7. Так, движение эмбрионов было впервые отмечено на 8-е сутки, в то время как при 2,6 %с — на 6-е. Вылупление при 29 %о началось через 12 сут. — на сутки позже, чем при 2,6 %. В опытах с соленостью 15 и 25 %с вылупление наблюдалось позже, чем при солености 2,6 %с, но раньше, чем при 29,0 %с Продолжительность вылупления также существенно различалась в опытах с различной соленостью. Наиболее коротким (3 сут.) оно было при 29,0%о, 4 сут. длилось вылупление при 15 и 25 %0, а наиболее длительным (7 сут.) было вылупление при 2,6%0. В последнем случае свободные эмбрионы дольше, чем в других опытах, оставались живыми после окончания вылупления (2 сут.). Раньше всего (через сутки после окончания вылупления) они погибали при 29,0 %о.
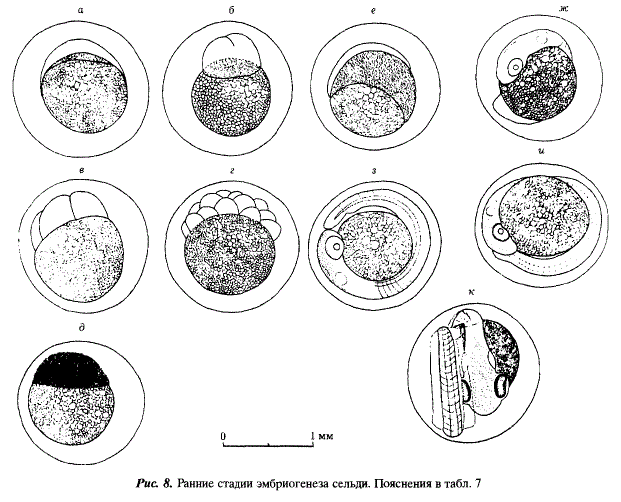
Таким образом, скорость дифференциации органов у эмбрионов лагунной сельди максимальна при развитии в воде с низкой соленостью. Это обстоятельство наряду с отмеченной выше наибольшей выживаемостью эмбрионов при низкой солености позволяет предположить, что последняя наиболее благоприятна для развития икры и зародышей лагунной сельди.
Интересно, что эмбрионы других групп сельди по-иному реагируют на пониженную соленость. Так, по данным И.Г. Фридлянд [1951], соленость ниже 4,65 %о является летальной для свободных эмбрионов сахалино-хоккайдской сельди. Икра охотской сельди также чувствительна к воздействию опреснения в периоды оплодотворения и выклева [Галкина, 1960]. По данным Б.В. Тюрина [1965], икра охотской сельди гибнет на нерестилищах, опресняемых речным стоком. Различия в отношении к солености являются одним из экологических механизмов репродуктивной изоляции между группами сельди, что позволяет существовать относительно малочисленным стадам лагунной сельди в тесном соседстве с высокочисленными стадами морских сельдей. В число таких механизмов входит также различное отношение к фотопериоду и температуре воды в период размножения. Лагунная сельдь, как уже говорилось выше, нерестует при большей продолжительности дня и при более высокой температуре воды, чем морская.

Наблюдения за размножением сельди в Ныйском заливе позволили установить причину низкого уровня ее численности, отмечаемого в течение последних лет и сказавшегося на величине уловов. Если с 1954 по 1966 г. уловы колебались от 7,7 до 32,9 тыс. ц, составляя в среднем 17,0 тыс. ц в год, то с 1967 по 1972 г. они не превышали 3,5 тыс. ц и в среднем равнялись 1,6 тыс. ц в год. Причиной столь значительного сокращения запасов, повидимому, явилось резкое уменьшение нерестового субстрата. Так, А.П. Веденский в 1935 г. видел, что "...90-95 % всей площади заливов восточного Сахалина густо заросли травами, и только узкие каналы (стоки речной воды) прорезают это сплошное травянистое море" [Веденский, 1950, с. 58]. То, что довелось наблюдать нам в 1972-1974 гг., "травянистое море" не напоминало. Это были редкие островки зостеры площадью от одного до нескольких метров и лишь изредка до нескольких десятков метров. По сведениям, полученным от местных рыбаков, уменьшение зарослей зостеры произошло в заливе в середине 60-х годов, что отчетливо совпадает со снижением уловов сельди. Интересно, что примерно в те же годы произошло сокращение полей зостеры в Белом море, также сопровождавшееся снижением численности сельди [Гемп, 1962]. Сокращение зарослей зостеры ухудшило условия размножения сельди не только из-за снижения емкости нерестилищ, но также в связи с уменьшением площади укрытий свободных эмбрионов и личинок. В начале 60-х годов, по данным В.Л. Андреева [1963], личинки сельди задерживались в заливе. В 1972-1973 гг. нам добыть их не удалось. В настоящее время они, скорее всего, сразу же после вылупления в значительном количестве выносятся из заливов сильными отливными течениями. Не исключено, что попадание в типично морские условия отрицательно сказывается на выживании свободных эмбрионов лагунной сельди.
