Зональные изменения состава населения и численности рыб в бассейне р. Начилова
Продольные смена сезонных ассамблей со специфическим составом нерестящихся и
нагуливающихся рыб была неоднократно показана на примере малых лососевых водотоков
Приморья (Богатов, 1994; Семенченко, 2001), Сахалина (Гриценко, 2002; Живоглядов, 2004),
Курильских островов (Сидоров, 2005), Японии (Nakano, Furukawa, 1990; Goto, Nakano, 1993) и
Северной Америки (Swales, 1987; Friesen, Ward, 1996). Камчатка является центром видового
разнообразия лососевых рыб и здесь существует значительное число речных систем, где одно-
временно обитает 9—10 видов этой группы, чего не отмечается в других регионах Северной
Пацифики. Очевидно, что в процессе совместного освоения речных бассейнов у лососевых
вырабатывались приспособления, способствующие снижению межвидовой конкуренции и
расхождению каждого вида по отдельным пространственным, нерестовым, пищевым и прочим
нишам. В результате, экосистемы камчатских рек способны устойчиво поддерживать существо-
вание популяций разных видов лососеобразных рыб.
В бассейне р. Начилова в течение всего года переход из одной речной зоны с характер-
ным диапазоном условий в другую сопровождается появлением новых видов рыб в уловах,
либо скачкообразным изменением во встречаемости уже имеющихся видов (табл. 6.1). Только
2 вида рыб (кижуч и мальма) обитают практически повсеместно и являются массовыми, при
этом доля кижуча вниз по течению растет, резко повышаясь в нижнем течении перед выходом в
устьевую зону, а доля мальмы, наоборот, сокращается. Микижа наиболее многочисленна
вблизи нерестового притока (р. Микочева) в зоне равнинных русел с перекатами. Численность
симы заметно сокращается при переходе из предгорий на равнину. Кунджа, хариус, 3-иглая и
амурская колюшки обитают только в зонах с равнинными руслами. Молодь нерки встречается
по всему бассейну эпизодически.
В летне-осенний период, когда на фоне относительно устойчивых абиотических условий
обостряется пространственно-пищевая конкуренция между разными размерно-видовыми груп-
пами рыб (Леванидов, 1981; The ecology…, 1986), в пределах каждой из выделенных зон
формируются сообщества со специфической структурой доминирования (табл. 6.7).
Распределение рыб по биотопам и укрытиям также характеризуется зональными разли-
чиями. На порожисто-водопадных руслах рыбы способны удерживаться лишь в эрозионных
ямах. На горных руслах без перекатов в июле и сентябре мальки лососей используют глайды
как транзитную область между отмелями и убежищами с пониженной мощностью потока. Пест-
рятки выходят на стрежень, где укрываются за выступающими камнями. На разветвленных
горных руслах с перекатами не заселены лишь открытые мелководные гребни перекатов, но
большая часть рыб держится в заломах и ямах-затонинах, либо выходит в пойменные протоки.
Водотоки с равнинными руслами характеризуются относительно равномерным распределением
рыб по акватории. В нижнем течении рр. Начилова и Микочева, отличающихся выровненным
рельефом дна, молодь отсутствует только на мелководных гребнях перекатов. Убежища не
играют ключевой роли при концентрации молоди, основные скопления которой отмечаются
на отмелях в тени нависающих крон деревьев и в затонинах за перекатами. В устьевой зоне
молодь и взрослые жилые рыбы концентрируются на стрежне плесовых лощин и в омутах.
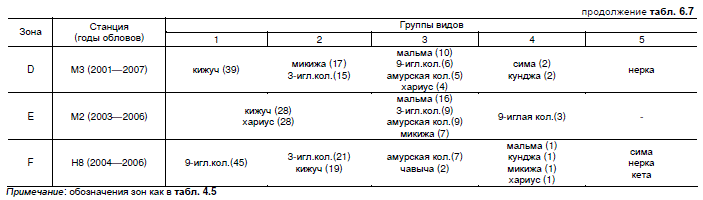
Средняя плотность заселения реки от истоков к устью в июле 2006 г. изменялась в 12
раз, биомасса — в 90 раз. Максимум плотности лососей (1,1—1,3 экз./м2) наблюдался в зоне
горных русел с перекатами, что связано с особо высоким разнообразием мест обитания и
нереста в русле и на пойме, а также в верхнем течении равнинного притока, что связано
с изоляцией крайне благоприятного для нагула локального участка равнинного русла отрезком
"торфяного канала". Вдоль равнинного русла главной реки средние значения плотности лосо-
сеобразных рыб плавно снижались с 0,8 до 0,5 экз./м2. Колюшек в нижнем течении реки было
102
Таблица 6.7. Средняя доля молоди и взрослых жилых рыб разных видов в июльских уловах станций полевых работ бассейна р. Начилова (%)
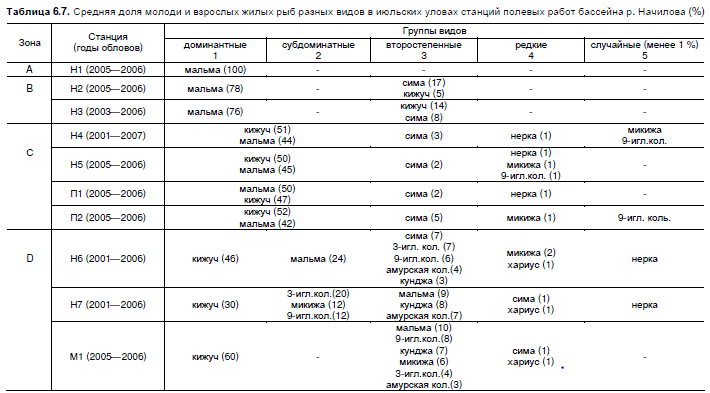
Примечание: обозначения зон как в табл. 4.5
104
не меньше лососей (плотности до 0,7 экз./м2), однако их биомасса в связи с мелкими размерами
не превышала 5 % от общей. Относительно высокая биомасса лососеобразных рыб (8—9 г/м2)
в равнинном поясе поддерживалась за счет высокой средней массы обитателей (14,1—110 г
против 8—10 г в предгорьях). В устьевой зоне с замедленным течением и однородными
условиями отмечена максимальная плотность колюшек — 2,2 экз./м2 (рис. 6.1).Рис. 6.1. Изменение средней июльской плотности обитания, экз./м2 (А) и биомассы, г/м2 (Б)
молоди и взрослых жилых рыб вниз по течению бассейна р. Начилова.
Показатели для: 1 — лососеобразных рыб в русле р. Начилова, 2 — колюшек в русле
р. Начилова, 3 — лососеобразных рыб в русле р. Микочева, 4 — колюшек в русле р. Микочева,
5 — лососевых рыб в русле руч. Предгорный.
Обозначения зон как в табл. 4.5.
Шестая станция в главном русле р. Начилова — устье р. Микочева.
105
Методом кластеризации доли (ln (% + 1)) видов в июльских уловах станций полевых
работ (табл. 6.7) в пределах бассейна выделяется 6 ассамблей рыб (рис. 6.2), каждая из
которых территориально приурочена к зоне бассейна со специфическим типом и морфологией
русел. Обособление ассамблей происходит на первых 4-х этапах ветвления дендрограммы,
уровень различий между ними составляет более 30 эвклидовых единиц. В наибольшей степени
от прочих в пространстве метрики Эвклида удалены ассамблеи молоди и взрослых жилых рыб
из истоков (станция Н1) и верхнего течения главной реки (Н2—Н3), ближе всего друг к другу
ассамблеи водотоков равнинных русел с перекатами (Н6—Н7—М1—М3) и без перекатов (М2).
Ветвление субкластеров, отражающее различия между населением станций в пределах одной
зоны, происходит после разделения населения на отдельные ассамблеи.
Рис. 6.2. Дендрограмма структуры рыбного населения бассейна
по встречаемости видов в уловах станций полевых работ.
Различия в структуре июльских ассамблей увеличиваются пропорционально длине
водного пути между ними. Значение коэффициента Шорыгина при сравнении населения
молоди и жилых рыб из соседних зон всегда ниже 1, в то время как между удаленными
зонами — больше 1 (табл. 6.8).
Таблица 6.8. Средние значения индекса ценотического различия Шорыгина между
ассамблеями, принадлежащими разным зонам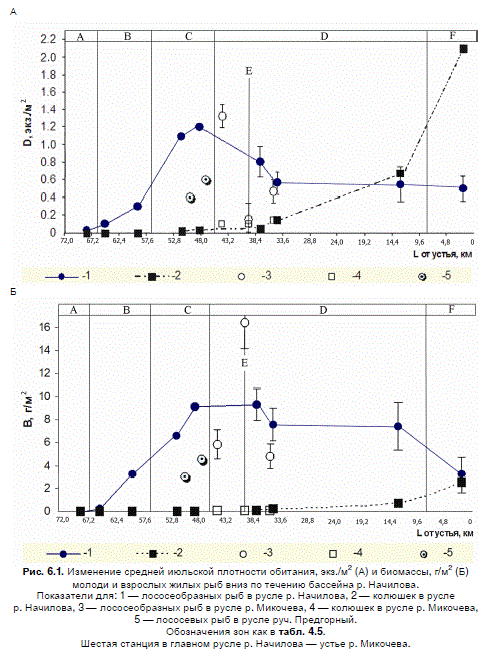
Примечание: обозначения зон как в табл. 4.5
Наибольшая сбалансированность (выравненность) структуры населения в июле харак-
терна для ассамблеи "болотного" участка, индекс доминирования здесь составляет 0,21,
нормированный индекс Шеннона равен 0,89. Доминирование незначительного числа видов в
ассамблее равнинных русел с перекатами выражено более отчетливо — средние значения
индексов составляют 0,28 и 0,72 соответственно. Несмотря на значительное разнообразие рыб
в уловах устьевой зоны, из-за сильного преобладания всего трех видов индекс выравненности
здесь ниже, чем на равнинных руслах, и составляет 0,60. Ассамблее устьевой зоны также свой-
ственна наибольшая энтропия (0,99). В предгорьях доминирование отчетливее проявляется в
ассамблее русел без перекатов — средний индекс равен 0,62, тогда как для населения раз-
ветвленных русел с перекатами показатель составляет 0,45. Вместе с этим для ассамблеи
неразветвленных русел также характерно более высокое значение нормированного индекса
106
выравненности видового состава Шеннона — 0,59 против 0,51, что связано с несбалансирован-
ностью ассамблеи разветвленных горных русел — энтропия ассамблеи выше, чем на
неразветвленных руслах в 2 раза. Ассамблея порожисто-водопадных истоков не сбалансиро-
вана (табл.6.9).
Таблица 6.9. Средние значения индексов доминирования (D), Шеннона (Н) и его
нормированной величины (ЕН), а также энтропийной оценки уровня сбалансированности (Q) для
рыбного населения станций полевых работ в июле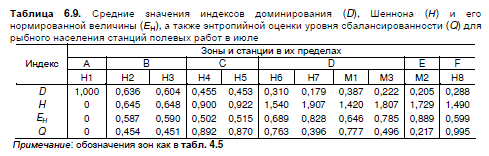
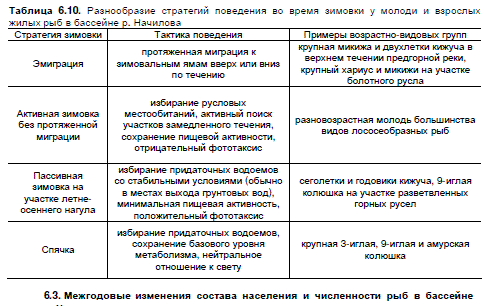
С наступлением зимы характер распределения молоди и взрослых рыб по речным зонам
меняется несущественно. Большая часть рыб остается зимовать на участках летне-осеннего
нагула. Анализируя закономерности выбора зимних местообитаний в бассейне р. Начилова,
можно заключить, что разнообразие стратегий зимовки рыб сводится к 4-м основным типам
(табл. 6.10).
Таблица 6.10. Разнообразие стратегий поведения во время зимовки у молоди и взрослых
жилых рыб в бассейне р. Начилова
Стратегия зимовки Тактика поведения Примеры возрастно-видовых групп
Эмиграция
протяженная миграция к
зимовальным ямам вверх или вниз
по течению
крупная микижа и двухлетки кижуча в
верхнем течении предгорной реки,
крупный хариус и микижи на участке болотного русла
Активная зимовка без протяженной миграции
избирание русловых местообитаний, активный поиск
участков замедленного течения,
сохранение пищевой активности,
отрицательный фототаксис
разновозрастная молодь большинства
видов лососеобразных рыб
Пассивная зимовка на участке летне-осеннего нагула
избирание придаточных водоемов
со стабильными условиями (обычно
в местах выхода грунтовых вод),
минимальная пищевая активность,
положительный фототаксис
сеголетки и годовики кижуча, 9-иглая
колюшка на участке разветвленных
горных русел
Спячка
избирание придаточных водоемов,
сохранение базового уровня
метаболизма, нейтральное
отношение к свету
крупная 3-иглая, 9-иглая и амурская
колюшка
Смотрите также
