Горные русла с перекатами
На разветвленные горные русла с перекатами на нерест поднимаются обе расы кижуча
и кеты, реофильная нерка, проходная и жилая мальма. Для проходной симы зона разветвлений
является транзитной на пути нерестовой миграции в верховья бассейна. Круглый год в районе
нерестилищ горных русел с перекатами держатся карликовые самцы мальмы. В летне-осенний
период из равнинных русел на нагул поднимаются "рекруты" (созревающие особи) и взрослые
особи микижи, а с горных русел без перекатов спускаются карликовые самцы симы. В поймен-
ных протоках и ключах воспроизводится 9-иглая колюшка.
Молодь рыб в сезонных уловах представлена личинками, мальками и пестрятками
мальмы, кижуча, симы и нерки, покатными мальками кеты (только в мае), а также личинками и
мальками 9-иглой колюшки (табл. 6.1).
Половодье (май-июнь). Подъем уровня воды на разветвленных руслах сопровождается
рассредоточением стока по временным пойменным протокам, в результате чего акватория уве-
личивается на порядок. Пестрятки мальмы возраста 1· — 2· и годовики симы, а также карлико-
вые половозрелые самцы мальмы (3·) в мае 2007 г. уже вышли из рукавов и основного русла во
временные пойменные протоки, где распределились по ямам и заломам. Часть крупной молоди
скапливается в заломах на затопленных оголовках островов главного русла. Плотность рыб в
укрытиях достигает 20—30 экз./м2. Вышедшие из грунта личинки мальмы начинают расселяться
по затопленным отмелям вблизи нерестилищ. Сеголетки кижуча и нерки подхватываются пото-
ком и уносятся вниз по течению с русловых нерестилищ. Часть молоди этих видов выносится в
пойменные протоки и надолго задерживается в зоне разветвлений. Кета, напротив, в районе
разветвлений не задерживается — вышедшие из бугров сеголетки ловятся только на стрежне.
Единичные особи 9-иглой колюшки держатся в пойменных протоках.
95
Меженно-паводочный период (июль-октябрь). Рассредоточение стока по рукавам и про-
токам сопровождается уменьшением мощности потока и ростом заселенности глайдов
с быстрым течением — крупные особи мальмы, симы и поднявшейся из равнинного пояса
микижи в июле выходят на открытые участки основного русла. Двухлетки мальмы и симы, а
также крупные сеголетки кижуча заселяют древесные заломы и промоины под берегами. Двух-
летки кижуча скапливаются в протоках и периферических заводях. Большая часть сеголеток
мальмы, кижуча и нерки, которые поздно вышли из грунта или попали во время половодья в
пойменные протоки, до поздней осени ловятся на отмелях горных русел. Во время кратковре-
менных паводков молодь лососей всех возрастов скапливается в заломах, заводях и
пойменных протоках.
В июле 2006 г. рыбное население было обильно даже на стрежне основного русла — в
среднем 0,2 экз./м². В отдельных рукавах присутствовали как участки глайдов с минимальной
плотностью обитания (< 0,05 экз./м2), так и обширные плесовые лощины с плотностью до
2 экз./м². Необитаемыми оставались лишь гребни перекатов. Количество молоди также сильно
различалось в рукавах разных размеров: на станции Н4 средняя плотность на 150-метровом
участке главного рукава составляла 0,25 экз./м², в одном из мелких вторичных рукавов — 0,6
экз./м², а в затененном пойменном ключе со слабым течением — почти 2,00 (± 0,4) экз./м2.
Плотность на отмелях и в укрытиях основного русла увеличивалась вниз по течению зоны раз-
ветвлений в 2—2,5 раза, а в древесных заломах — в 5 раз. В отдельных случаях плотность в
заломах достигала 13 экз./м2 (станция Н4) — 15 (Н5) экз./м2. В эрозионных ямах и мелких пой-
менных протоках количество молоди лососей при движении вниз по течению менялось слабо
(табл. 6.3).
Таблица 6.3. Характеристика ассамблеи молоди и взрослых жилых рыб на горных руслах
с перекатами в июле
Средняя плотность заселения
(среднее ± ошибка), экз./м2 дна
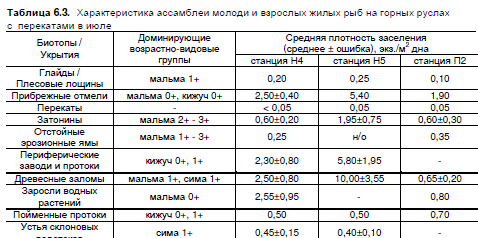
Средняя июльская плотность обитания в 2006 г. в русловых местообитаниях станции Н4
составила 0,70 экз./м2, в пойменных — 1,5 экз./м2; эти же показатели для станции Н5 — 0,85
и 2,1 экз./м2. Общая плотность заселения реки лососями и колюшками (биотопы всех типов) в
период исследований на станции Н4 составила 1,10 экз./м2, биомасса — 6,8 г/м2; на станции
Н5 — 1,25 экз./м2 и 9,2 г/м2 соответственно. Средняя масса 1 экз. лососевых рыб в июле коле-
балась от 7,6 (Н4) до 7,7 (Н5) г; средняя масса 1 экз. колюшек — 1,1 г.
Плотность заселения среднего и нижнего течения руч. Предгорный в конце июля 2006 г.
была ниже соответствующих показателей в главном водотоке зоны разветвлений. Средняя
плотность лососевых рыб на участке в 4-х км выше устья ручья (станция П1) — 0,4 экз./м2, рыба
была распределена по акватории равномернее, чем в главной реке. В 1 км выше устья (П2)
плотность составила 0,6 экз./м2, при этом в некоторых пойменных протоках показатель достигал
10—15 экз./м2.
В конце августа на глайдах и прибрежных отмелях основного русла и рукавов начина-
ется совместный нерест проходной и жилой мальмы, а в затонинах за перекатами, под берегом
— нерест ранней кеты. С сентября эрозионные ямы, а также глубокие прибрежные участки пле-
96
совых лощин и глайдов основного русла занимают производители раннего кижуча, в пойменные
протоки, култуки и лососевые ключи заходит поздняя кета и нерка.
Устойчивая межень (ноябрь-апрель). В начале декабря 2006 г. в уловах встречалась
молодь мальмы, симы и кижуча, а также 9-иглая колюшка всех возрастов. В течение зимы для
водотоков зоны характерно чередование периодов активного намерзания поверхностного
и донного льда, ледостава, когда образуются широкие забереги и стрежень засыпает снегом, а
также периодов, когда в результате оттепелей поверхностный и глубинный лед разрушается
и уносится вниз по течению. Шугоходы возможны и в период ледостава. Двигающаяся в толще
воды шуга забивает рукава и пойменные протоки, перегораживает основное русло на перека-
тах, меняет направление потока в плесовых лощинах. Образование ледяных заторов приводит
к отчуждению части местообитаний и смерти молоди лососей. Основным фактором, опреде-
ляющим выживаемость рыб в течение цикла ледовых явлений, остается доступность укрытий
(Prowse, 1995). Для малых водотоков Северной Америки показано, что в результате мощных
шугоходов смертность зимующих в руслах рыб может достигать 40—60 % (Bustard, 1986;
Сunjak, 1996), в то время как в незамерзающих водотоках молодь почти не погибает (Huusko et
al., 2007). В р. Начилова 11 декабря 2006 г. в шуге, прибитой к берегу, были обнаружены мерт-
вые особи мальмы в возрасте 1+ и 2+.
В отличие от вышележащей зоны со стабильным ледовым режимом, где все рыбы
зимуют в схожих местообитаниях, динамичность зимних условий разветвленных русел
предопределяет наличие нескольких типов стратегии зимовки молоди и взрослых жилых рыб
(Сunjak, 1996; Enders et al., 2007). Крупная микижа и двухлетки кижуча, которые в течение лета
и осени держались на глубоких участках русла и в заводях, в начале зимы мигрируют на рав-
нинные русла с перекатами. Молодь мальмы и симы далеко не мигрирует, в период оттепелей
она отстаивается на оконечностях подрезанных побочнем отмелей — на границе мелководий
и глайдов, в устьях пойменных проток и в заводях на границе заберегов и открытой воды. Во
время ледообразования большинство молоди укрывается позади заломов из древесного топ-
ляка, которые препятствуют продвижению льда (Bustard, Narver, 1975; Сunjak, 1996; Harper,
Farag, 2004), а сеголетки мальмы — под камнями. Сеголетки кижуча покидают заводи и отмели
основного русла и рукавов и скапливаются в пойменных ключах и лимнокренах на выходах
грунтовых вод, которые четко фиксируются по местам открытой воды в промерзших и засыпан-
ных снегом русловых ответвлениях. Летом в холодноводные ключи и лимнокрены (температура
воды 5—7 ºС) рыба заходит только в случае стрессово высоких температур воды (выше 18—
20 ºС) в главном русле (Snucins, Gunn, 1995), зимой здесь образуются одновидовые стаи пест-
ряток кижуча численностью до нескольких сотен штук. Значение незамерзающих пойменных
биотопов в зимовке молоди лососей в северной Канаде подтверждается наличием пропорцио-
нальной связи между выживаемостью кижуча, чавычи, гольца (S. fontinalis) и микижи и
количеством ключей на длину малой реки (Bustard, 1986; Smith, Griffith, 1994; Сunjak, 1996).
Колюшка всех возрастов зимой ведет себя пассивно, держась в глубоких пойменных протоках и
заводях подо льдом. Последние промерзают почти до дна, однако для выживания мелкой рыбы
в состоянии спячки достаточно пространства в 3—5 см между льдом и дном (Berg, 1994).
Смотрите также
