Характеристика кормовой базы молоди лососевых рыб в бассейне Начилова
Установлено, что обеспеченность молоди лососей пищей в реках влияет на ее выжи-
ваемость и рост не только во время пресноводного нагула, но и на ранних этапах морского
(Леванидов, 1969). В камчатских реках истинный зоопланктон отсутствует, и молодь лососей
питается главным образом макрозообентосом. Однако, по определению Е.В. Боруцкого (1960)
материалы бентосных проб дают представление о кормовых ресурсах рек, а кормовая база ―
"это та часть кормовых ресурсов, которая может быть использована в качестве пищи наличным
видовым и возрастным составом ихтиофауны при условии существующих в водоеме биотиче-
ских и абиотических взаимоотношений". В лососевых реках с быстрым течением и плотными
гравийно-галечными грунтами кормовая база молоди существенно отличается от кормовых
ресурсов, поскольку доступность донных беспозвоночных, населяющих, как правило, нижнюю
поверхность валунов и щели между галькой, ограничена (DeWalt, Stewart, 1995; Ward, 1992).
Молодь способна утилизировать не более 30—50 % продукции макрозообентоса (Waters, 1988).
Существенную роль в повышении доступности донных беспозвоночных для рыб играет
дрифт, о его роли как кормовой базы молоди и жилых лососевых свидетельствует сходство
спектров и ритмов питания рыб со структурой и динамикой дрифта в течение суток и в сезон-
ном аспекте (Фроленко, 1970; Чебанова, 1983б, 2002; Ringler, 1985; Kreivi et al., 1999; Amundsen
et al., 2000; Giroux et al., 2000). Установлено, что от обилия дрифта зависят интенсивность
питания и рост молоди, особенно территориальной (Lister, Genoe, 1970; McNicol et al., 1985;
Wilzbach et al., 1986; Brannas, Alanara, 1994; Nacano, 1995, Sommer et al., 2001). Например,
сравнительный анализ популяций микижи нескольких небольших рек в Каскадных горах Север-
ной Америки показал прямую связь темпа роста молоди с количеством дрифта в соответст-
вующий период (Murphy et al., 1981). Большинство исследователей придерживаются мнения,
что само территориальное поведение лососевых — это конкуренция за микробиотопы, удобные
для питания дрифтом (Grant, 1990; Gotceitas, Godin, 1992; Mikheev et al., 1994; Nacano, 1995;
Adams, Huntingford, 1996). Соответственно, с плотностью дрифта связаны площади индивиду-
альных кормовых территорий, и, в конечном счете, плотность заселения участков реки
(Chapman, 1966). Кроме того, различия в обеспеченности пищей на разных участках приводят к
дифференциальному росту территориальной молоди (Chapman, 1962). Питание эпибентосом
преобладает только у личинок и мелких сеголеток, обитающих в небольших мелких заводях
вдоль береговой линии (Mundie,1969; Grant, Noakes,1987; Hubert, Rhodes, 1992) и у молоди в
мутных реках (Tippets, Moyle, 1978). Таким образом, в лососевых реках обеспеченность молоди
рыб легкодоступным кормом напрямую зависит не только от обилия донных беспозвоночных на
дне, но и от их миграционной активности, т.е. обилия в толще воды (Allan, 1981, 1995; Rader,
McArthur, 1995).
Рассматривая бентос и дрифт как кормовую базу молоди лососей необходимо учиты-
вать размерный состав донных беспозвоночных, поскольку анализ обширного материала по
питанию различных видов лососевых в камчатских реках показал, что личинки ручейников
O. unicolor, B. americanus, веснянок сем. Perlodidae и типулид крупнее 15—17 мм, а также мол-
люски с диаметром раковины более 4 мм не встречаются в желудках молоди, даже у крупных
134
двухлеток кижуча и чавычи. Соответственно, биомассу собственно "кормового" бентоса и
дрифта следует рассчитывать без учета этих крупных организмов.
Роль отдельных таксонов донных беспозвоночных в питании молоди неравнозначна. Как
показали многочисленные трофологические исследования, в реках основным кормом всех
видов лососевых рыб служат амфибиотические насекомые, в первую очередь доминирующие
по численности хирономиды (Леванидова, 1982; Николаева 1968; Williams, 1981; Cunjak, 1992).
Согласно детальной классификации Рэдера (Rader, 1997), наиболее доступными и важными
кормовыми объектами являются многочисленные, активно участвующие в дрифте хирономиды,
мошки, поденки семейств Baetidae, Heptageniidae, родов Ephemerella, Drunella, Paraleptophlebia,
Ameletus, а также бокоплавы. К второстепенным кормовым объектам автор отнес не склонных к
активным миграциям веснянок, в том числе родов Alloperla, Suwallia, Capnia, поденок
сем. Ephemeridae, ручейников сем. Glossosomatidae, некоторых двукрылых, а также олигохет,
планарий и прочих беспозвоночных, обитающих в отложениях мелкого гравия, песка и ила.
Следует отметить, что в период массового вылета амфибиотических насекомых молодь
лососей активно питается не только их личинками, но и имаго. Обеспеченность рыб этими лег-
кодоступными кормовыми объектами на каждом участке реки в значительной мере зависит от
обилия амфибиотических насекомых в бентосе. Рыбы схватывают имаго в толще воды после
их смерти, с поверхности воды во время откладки яиц или прямо в воздухе во время роения (в
прыжке).Кроме того, в момент массового нереста тихоокеанских лососей молодь рыб способна
переходить на активное питание вымываемой с нерестилищ икрой (Гриценко, 2002;
Живоглядов, 2004; Chaston, 1969). Поздней осенью и в начале лета до трети пищи молоди ло-
сосей может составлять мышечная ткань разлагающейся "сненки" (Введенская, Травина, 2008;
Кирилова, 2008). Подросшие лососи переходят к факультативному хищничеству.
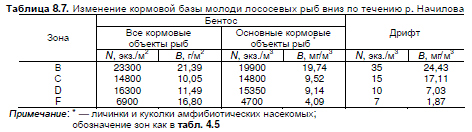
Характеризуя кормовую базу р. Начилова, количественные характеристики бентоса и
дрифта рассчитывали без учета крупных ручейников и моллюсков, а также выделяли группу
амфибиотических насекомых, как основных кормовых объектов молоди лососей. От верховьев
к устью реки количественные показатели "кормового" бентоса снижаются (табл. 8.7). Макси-
мальная численность и биомасса кормовых объектов (23 тыс. экз./м2 и 21 г/м2), в том числе
амфибиотических насекомых (20 тыс. экз./м2 20 г/м2), наблюдаются в зоне горного русла без
перекатов. В зоне разветвлений и в равнинном поясе показатели обилия "кормового" бентоса
практически одинаковы — 15―16 тыс. экз./м2 и 10―11 г/м2 (амфибиотические насекомые —
15 тыс. экз./м2 и 9 г/м2). На устьевом участке реки общая численность кормовых объектов сни-
жается до 7 тыс. экз./м2, а биомасса, благодаря крупным олигохетам, увеличивается до 17 г/м2.
При этом амфибиотических насекомых на заиленном русле мало ― 5 тыс. экз./м2 и 4 г/м2.
Тенденция снижения обеспеченности молоди кормом вдоль русла реки еще отчетливее про-
слеживается на изменении количественных показателей "кормового" дрифта. Численность
легкодоступных в толще воды беспозвоночных в зонах горных русел без перекатов и с перека-
тами, равнинных русел с перекатами и на устьевом участке составляет 35, 15, 10 и 7 экз./м3,
биомасса — 24, 17, 7 и 2 мг/м3 соответственно. Высокую биомассу "кормового" дрифта в пред-
горном поясе обеспечивают миграции относительно крупных поденок старших возрастов.
Таблица 8.7. Изменение кормовой базы молоди лососевых рыб вниз по течению р. Начилова
Примечание: * — личинки и куколки амфибиотических насекомых;
обозначение зон как в табл. 4.5
В нижнем течении равнинной реки Микочева численность и биомасса "кормового" бен-
тоса составляют в среднем 18 тыс. экз./м2 и 9,35 г/м2, амфибиотических насекомых ― 17 тыс.
экз./м2 и 7,72 г/м2. Без учета массовых на этом участке реки ручейников сем. Glossosomatidae
(A. inaequispinosus), относящихся по классификации Рэдера к второстепенным кормовым объ-
ектам молоди лососей, численность и биомасса амфибионтов снижаются до 16 тыс. экз./м2 и
5,70 г/м2. Судя по размерному составу мигрантов, весь дрифт в р. Микочева можно считать
"кормовым", однако его плотность и биомасса невелики ― 7 экз./м3 и 3,26 мг/м3.
Смотрите также
