Долгопериодные изменения биологических показателей осенней кеты
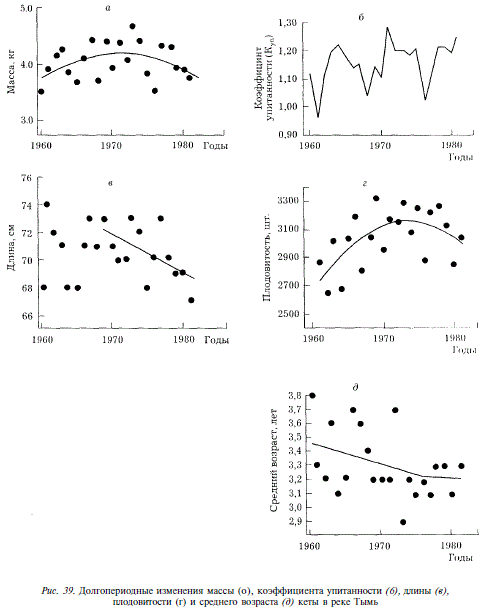
Долгопериодные изменения биологических показателей кеты мы рассмотрим на примере стада Тыми, по которому собраны наиболее систематические данные за длительный ряд лет. С 1960 по 1981 г. средняя длина кеты Тыми колебалась от 67 до 74 см, средняя масса — от 3,53 до 4,70 кг. Четкой периодичности колебаний этих показателей в рассматриваемый отрезок времени не наблюдалось. Уменьшение или увеличение длины, как правило, сопровождаются соответствующими изменениями массы. Исключение составляют лишь 1962, 1963, 1971 и 1978 гг., когда направленность изменений длины и массы была противоположной (рис. 39,а,в). Начиная с 1969 г., намечается тенденция уменьшения длины кеты. По иному изменялась масса рыб: с 1960 по 1973 г. она увеличивалась. После 1973 г. масса начала уменьшаться. Эти изменения можно описать уравнением параболы у = 3,660 + 0,08993 х 0,00368 х2.
В изменениях упитанности (К,) отмечалось два периода, включающих подъем и спад продолжительностью 7 и 8 лет (см. рис. 39,6). В изменениях плодовитости также, как в изменениях массы, отмечаются подъем и спад (см. рис. 39,г). Ход изменений плодовитости описывается уравнением у = 2667 + 74,34 х — 2,814 х2. Перегиб кривой приходится на 1972-1973 гг., т.е. практически совпадает с перегибом кривой изменений массы.
Средний возраст нерестового стада с 1960 до 1981 г. колебался от 2,9 до 3,8 лет (при вычислении среднего возраста мы использовали число целых лет, прирост года наблюдений не учитывался). На этом отрезке времени выделяются два периода: первый — с 1960 по 1973 г., когда межгодовые колебания были значительны, и второй — с 1974 по 1981 г., во время которого средний возраст был относительно постоянен и находился на низком уровне (см. рис. 39,д).
Ход изменений среднего возраста кеты описывается уравнением
У= 3,466 -0,004353 x
У. Риккер [Ricker, 1980], анализировавший многолетние изменения размеров и возраста кеты Аляски и Британской Колумбии в период с 1951 по 1975 г., отмечает уменьшение ее длины и массы при одновременном незначительном увеличении среднего возраста. По его мнению, это результат селективного изъятия дрифтерным промыслом наиболее быстрорастущих и раносозревающих особей. Постоянное изъятие их из генофонда должно сказываться на всех возрастных группах, так как между ними происходит скрещивание. Однако воздействие промысла на возрастную и размерную структуру кеты американских стад существенно отличается от воздействия на азиатские стада. Американский промысел изымает преимущественно половозрелую кету в морских прибрежьях, на подходе к нерестовым рекам. Азиатская кета облавливается японским флотом на обширных участках открытого океана, при этом неполовозрелые особи составляют значительную часть уловов. В этих условиях кета той или иной генерации неоднократно на протяжении жизни попадает под воздействие морского промысла. При этом у рыб, созревающих в старшем возрасте, такие контакты бывают чаще и продолжительнее. Естественно, что быстрорастущие и раносозревающие особи вылавливаются морским промыслом в первую очередь.
Таким образом, кета азиатских стад, в отличие от американской, подвержена двум противоположно направленным селективным воздействиям промысла. Давление отбора, изымающего особей старших возрастов, видимо, преобладает, поскольку в целом для кеты Тыми характерно измельчание и омоложение. В то же время относительная стабилизация среднего возраста в течение последних 8 лет наблюдений говорит о действии и противоположно направленного отбора.
Однако селективное воздействие морского промысла при всей его важности не может являться единственным фактором, определяющим облик азиатских стад кеты. Естественный ход природных процессов, годовые особенности условий нагула также должны сказываться на параметрах популяций. Судить об их воздействии мы можем лишь па основании косвенных показателей. С этой целью мы попытались проанализировать зависимость средних длины и массы нерестовых стад кеты от их среднего возраста. Оказалось, что эти характеристики связаны параболической зависимостью (рис. 40). Восходящая ветвь параболы, по нашему мнению, определяется благоприятными условиями нагула в океане, когда увеличение в нерестовом стаде доли особей старших возрастов сопровождается увеличением средних размеров производителей. Нисходящая ветвь обусловлена годами с неблагоприятными условиями нагула, при которых рыба растет медленнее и созревает в более старшем возрасте. После нагула в неблагоприятных условиях на нерестилища приходят мелкие и относительно старые производители.
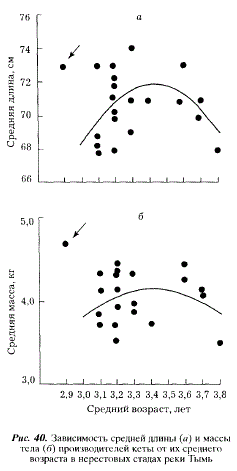
Особняком в наших данных стоит точка с абсциссой 2,9 (1973 г.), помеченная на рис. 40 стрелкой. Поскольку она находится вне основной группировки точек и не подчиняется общей закономерности их распределения, мы не использовали ее в расчетах. Однако положение этой точки не нарушает логики наших рассуждений. Видимо, раннее созревание и исключительно быстрый рост кеты происходят в редких случаях в годы с очень благоприятными условиями нагула, после которых на нерестилищах появляются крупные и относительно молодые производители.
