Воспроизводство запасов
Нерест кеты в бассейне Тыми начинается обычно в последней декаде августа. Примерно 80 % производителей нерестятся в период с последней декады сентября до конца октября. Последние пары производителей встречаются на нерестилищах в течение декабря.
От начала к концу нереста среднесуточная температура воды в основном русле Тыми снижается с 12 до 0 °С. Температура воды в ключах и лимнокренах в конце нереста бывает выше: в ноябре 2-4°С, в декабре 1,2-2,5°С. Нерест кеты проходит в период осеннего повышения уровня воды, которое начинается обычно со второй половины сентября, после выпадения осенних дождей. В отдельные годы осенние паводки превышают паводки весенние. Так, в 1961 и 1965 гг. сентябрьские расходы воды достигали в верховьях реки соответственно 136 и 283 м3 /с, в то время как расходы воды во время весенних паводков — лишь 126 и 144 м3/с. К началу паводков на нерестилища проходит 10-15 % производителей, а в целом влияние паводков, длящихся до середины октября, ощущают на себе 70-90 % нерестящихся рыб.
Нерестилища кеты внутренней дельты Тыми и притоков верхнего течения обильно снабжаются теплыми грунтовыми водами, и обширные поверхности нерестовых водоемов не замерзают даже в самые суровые морозы. Лед образуется, как правило, лишь у берегов и на ямах. Температура воды на этих незамерзающих нерестилищах в декабре — январе колеблется от 0,5 до 1,5°С. В марте она повышается до 1,0-2,1°С, к середине апреля — до 3,5-5,0 °С.
Поскольку нерест основной массы кеты происходит при повышенном уровне воды, после его падения в зимнюю межень значительная часть нерестовых бугров оказывается выше уреза воды. Выходы грунтовых вод и высокий снежный покров надежно защищают эмбрионы в этих буграх. Промерзание бугров кеты в бассейне Тыми невелико. Однако после сильных осенних паводков оно бывает выше, чем в годы с низкими уровнями воды. Так, зимой 1962/63 г., во время которой поддерживался нормальный уровень, промерзло 7-8 % нерестилищ, где погибла икра от 12 тыс. производителей, а зимой 1961 — 1962 г. после спада паводковых вод промерзло 12-14 % нерестилищ с икрой от нереста 75 тыс. рыб [Гриценко, 1967].
О глубинах, предпочитаемых кетой .при нересте, можно судить с известной осторожностью, так как в течение нереста уровень воды сильно меняется. Оптимальными, видимо, являются глубины от 30 до 80 см. По мере снижения уровня воды часть бугров оказывается выше уреза воды. Из тех бугров, которые остались под водой, 90 % находятся на глубине 10-40 см, 50 % — на глубине 12-22 см. Скорость течения во время нереста 0,05-1,0 м/с. Минимальные скорости течения отмечаются в лимнокренах. Скорость течения на нерестилищах в феврале не превышает 0,7 м/с, составляя в среднем 0,3 м/с. В лимнокренах и ключах поверхностное течение часто отсутствует. Средняя высота бугров 14 см при колебаниях от 2 до 24 см. Бугры имеют разную форму, чаще всего это эллипсы, вытянутые по течению или поперек течения. Нередки бугры в форме серпа, обращенные выпуклой стороной вниз по течению. По данным Ф.Н. Рухлова [1972], средняя длина бугров кеты в Тыми 1,39 м при колебаниях от 0,9 до 2,6 м; средняя ширина 1,16 м при колебаниях от 0,8 до 1,8 м.
Верхняя граница расположения икры в буграх находится на глубине 6-63 см (в среднем 25 см), нижняя граница — на глубине 14-80 см (в среднем 38 см). Средняя толщина слоя икры в буграх 13 см при колебаниях от 3 до 65 см. Личинки накануне выхода из бугров обнаруживаются на меньшей глубине — верхняя граница их нахождения в 8 20 см от поверхности. Около 90 % вскрытых бугров содержали от 200 до 1400 икринок, 75 % —300-900 икринок и 50 % — 400-800 икринок. Икра в буграх чаще всего располагается двумя хорошо выраженными гнездами. Несколько реже таких гнезд бывает три или одно. Примерно в 15-20 % бугров гнезда не выражены, а икра в них расположена дисперсно по всей площади.
В среднем в одном бугре выживает 333 личинки, что составляет 10,9 % от средней абсолютной плодовитости (табл. 24). Основные потери икры приходятся на период нереста, во время которого в бугры попадает лишь 23,3 % от средней абсолютной плодовитости, остальная же икра уносится течением и поедается хищниками. В период между осенним и весенним вскрытиями бугров, длящийся около 4 мес, примерно 30 % отложенной икры исчезает, полностью разлагаясь, или поедается беспозвоночными. Потери за весь период пребывания в грунте составляют в среднем около 50 % от отложенной икры.
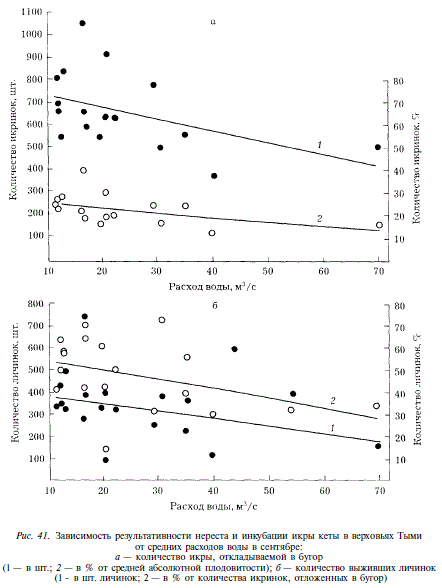
Рассмотрим, как зависит эффективность нереста и выживания эмбрионов кеты от воздействия различных факторов. Количество икры, откладываемой в среднем в один бугор, не зависит от величины плодовитости самок (и = 19; г = 0,016) (табл. 25). Увеличение расходов воды в сентябре приводит к уменьшению как абсолютного, так и относительного (процент от абсолютной плодовитости) количества икры в буграх за счет выноса икринок во время нереста (рис. 41). Связь между этими показателями, видимо, более тесная, чем удалось установить. Дело в том, что практически бывает трудно обнаружить бугры, устроенные при очень высоком уровне. При определении зависимости мы не включили в расчет два года с экстремально высокими паводками, так как в эти годы вскрывались бугры, устроенные лососями уже после прохождения паводковых вод. Расходы воды в октябре меньше влияют на количество икры, откладываемой в бугры. Связь между этими показателями отрицательная, но коэффициент корреляции ниже достоверного уровня.
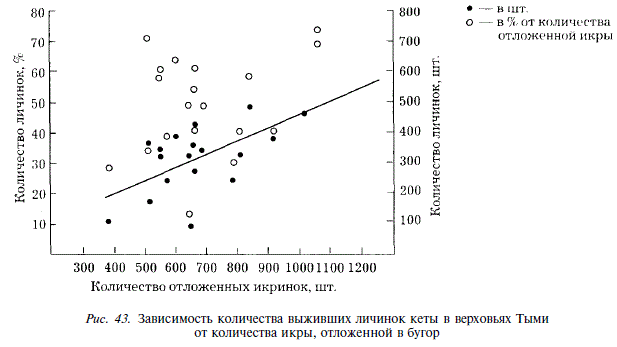
Среднее количество икры в одном бугре связано с численностью производителей на нерестилищах параболической зависимостью (рис. 42,а). Такой вид зависимости оказался для нас несколько неожиданным. Напомним, что у горбуши ключа Холодный эти показатели связаны отрицательной линейной зависимостью. Характер связи численности производителей кеты с количеством икры, откладываемой в бугор, видимо, обусловлен фактором беспокойства, которое испытывает кета в таком густо населенном районе, как верховья Тыми. Беспокойство тем выше, чем ниже численность рыб, но. начиная с какого-то момента, уменьшение количества икры в буграх определяется у кеты численностью производителей. Количество выживших эмбрионов связано прямой линейной зависимостью с количеством икры, отложенной в бугор (г = 0,61) (рис. 43). Вместе с тем, процент выживания молоди от количества отложенной икры не зависит. Увеличение расходов воды в сентябре приводит к уменьшению абсолютного количества выживших личинок, и это естественно, так как сентябрьские паводки обусловливают уменьшение количества икры, откладываемой в бугры, а последнее прямо связано с количеством выживших личинок. Однако сентябрьские паводки вызывают также уменьшение процента выживаемости эмбрионов (г = -0,408) (см. рис. 41,б). Механизм этого явления нельзя объяснить однозначно: увеличение расходов воды во время нереста может быть причиной повышенного травмирования эмбрионов. Так, по данным У. Мак Нейла [McNeil, 1966], движение грунта в период высокой воды вызывает гибель 50-90 % эмбрионов кеты и горбуши, находящихся в буграх.

Во время очень высоких паводков гольцы перестают выполнять на нерестилищах функции санитаров, поедающих потерянную при нересте икру [Гриценко, 1969в]. При высоких скоростях течения гольцы укрываются в тихих местах. Икра же, уносимая течением, будучи намытой в грунт нерестилищ и разлагаясь там, может вызвать повышенный отход в буграх.
Связь между величиной расхода воды в октябре, январе и выживаемостью личинок отсутствует. Благодаря обильному выходу грунтовых вод и высокому снеговому покрову эмбрионы кеты р. Тымь благополучно выживают даже при низких зимних уровнях воды в реке. Ситуация, существующая в этой реке, отличается от таковой в реках Камчатки, где выживаемость кеты зависит от количества осадков, выпадающих в январе [Николаева, 1980]. При увеличении осадков усиливается интенсивность грунтового стока и уменьшается промерзаемость бугров.
Численность производителей на нерестилищах связана с выживаемостью личинок параболической зависимостью (см. рис. 42,6). При слабом заполнении нерестилищ пониженная выживаемость, видимо, обусловлена недостаточной очисткой нерестилищ от ила в процессе устройства бугров, недостаточным их рыхлением. Пониженная выживаемость личинок при высокой численности производителей вызывается перекопкой ранее устроенных бугров, большими потерями икры при нересте, поскольку увеличивается попадание мертвой икры в гнезда, вследствие чего возрастает вероятность поражения живой икры сапролегнией.
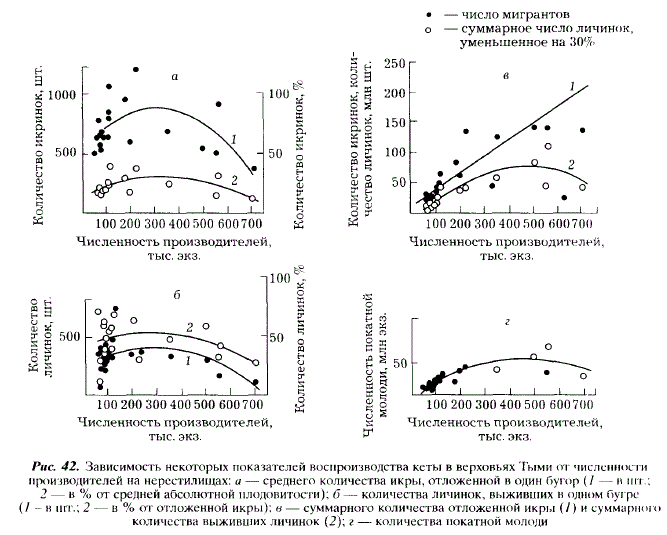
Сказанное выше относится к результативности нереста одной пары производителей, а суммарное количество икры, откладываемой нерестовым стадом, линейно увеличивается с увеличением численности производителей. Суммарное выживание личинок в буграх, возрастая по мере увеличения численности производителей от 450-500 тыс. экз., начинает снижаться после превышения этой величины. Аналогичная зависимость существует между численностью производителей и количеством покатной молоди: наибольшее число молоди обеспечивается нерестом 450-500 тыс. производителей (см. рис. 42,г). Рассмотренная зависимость обусловлена тем, что нерест части производителей кеты в годы высокой численности происходит на участках реки, не пригодных для размножения этого вида, и неизбежно приводит к снижению количества покатной молоди.
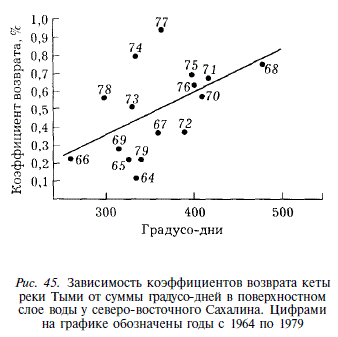
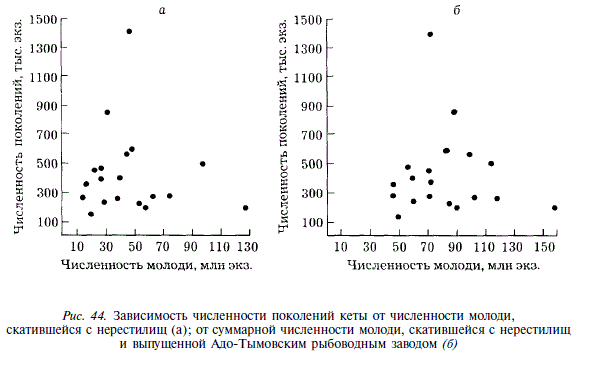
Несмотря на то, что молодь кеты скатывается в море при более крупных размерах, чем горбуша, а значительная ее часть начинает питаться еще в реке, связь между численностью скатившейся молоди и численностью вернувшихся поколений отсутствует. Нет даже такой слабой связи, какая наблюдалась нами у горбуши. Коэффициент корреляции между величиной ската с нерестилищ и численностью поколений — 0,086, между суммарным скатом молоди с нерестилищ и с рыбоводного завода и численностью поколений — 0,056 (рис. 44). Такая ситуация возможна в том случае, если численность поколений кеты формируется в прибрежных участках моря под влиянием факторов среды, действие которых имеет случайный характер. К подобным выводам пришел A.M. Каев [1983], исследовавший воспроизводство кеты острова Итуруп. Наши данные подтвердили его точку зрения. Сопоставление коэффициентов возврата кеты с термическими условиями нагула в прибрежье (в качестве их показателя мы используем сумму градусодней в июне — июле в поверхностном слое воды) показало четкую тенденцию увеличения коэффициентов возврата с увеличением суммы тепла (рис. 45). Связь описывается уравнением у = -0,358 + 0,00238 х (г = 0,52; Р = 0,05).
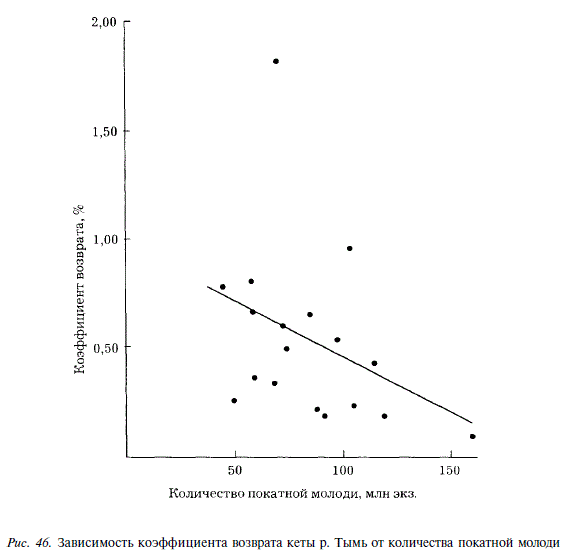
Низкие температуры воды могут действовать на молодь непосредственно, вызывая у нее термический шок при попадании в море, могут, а также влиять опосредованно, через замедление развития кормового планктона и ухудшение условий роста. Кроме того, молодь тымской кеты подвержена в прибрежье моря действию факторов смертности, зависящих от плотности, так как величина Кв уменьшается с увеличением количества покатников. Зависимость описывается уравнением у = 0,971 0,0051Л:; Г = 0,35 (рис. 46).