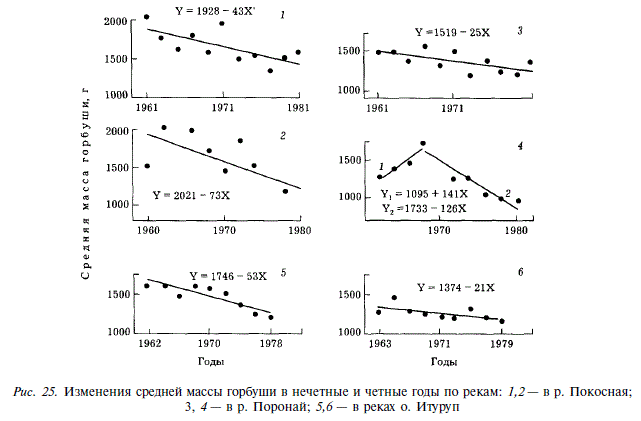
Долгопериодные изменения размеров горбуши
В течение последних двух десятилетий наблюдается измельчание горбуши как на япономорском, так и на охотоморском побережьях Сахалина (рис. 25). Одновременно отмечается измельчание горбуши Южных Курил [Ефанов, Чупахин, 1982]. Явление это подобно измельчанию горбуши Британской Колумбии и юго-восточной Аляски [Ricker et al., 1978]. Поскольку основными причинами, вызывающими уменьшение размеров горбуши, считают влияние селективного промысла и ограниченность кормовых ресурсов, рассмотрим возможное действие каждой из них.
Азиатская горбуша, в том числе и горбуша Сахалина, начиная со второй половины 50-х годов, находится под мощным воздействием японского морского промысла. Наиболее интенсивным это влияние было в 60-е годы, в результате чего в ряде районов численность горбуши сократилась настолько, что она утратила свое промысловое значение (например, западная Камчатка). Благодаря мерам международного регулирования вылов горбуши в открытом море стал постепенно сокращаться и с 40-70 тыс. т в 60-е годы снизился к концу 70-х годов до 17-30 тыс. т. С середины 60-х годов началось увеличение численности горбуши Южных Курил и охотоморского побережья Сахалина, а начиная с середины 70-х годов — горбуши западной Камчатки. Одновременно с увеличением численности горбуши наблюдается ее измельчание (по крайней мере, рыб, воспроизводящихся на восточном Сахалине, — охотоморской летней и охотоморской осенней популяций. На западном побережье Сахалина ситуация была другой. Воздействие японского морского промысла на япономорскую горбушу все время оставалось высоким. Численность же япономорской горбуши после подъема, отмечавшегося в конце 60-х — начале 70-х годов, находилась до 1989 г. на низком уровне. В первую очередь это относится к той части япономорской популяции, которая воспроизводится в реках западного Сахалина, где численность заходящих на нерест рыб резко сократилась после 1969 г.
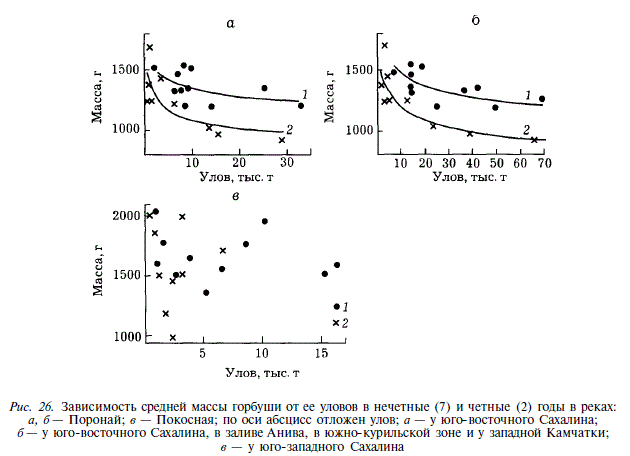
Масса горбуши восточного Сахалина и залива Анива уменьшалась по мере увеличения численности популяций, причем у поколений четных лет это изменение происходило быстрее. В качестве примера рассмотрим горбушу реки Поронай (рис. 26). Зависимость средней массы горбуши в этой реке от численности (в качестве показателя численности мы использовали величину уловов в северной части залива Терпения) описывается для нечетных лет уравнением у-1841 х"0'066; для четных лет — уравнением у = 1658 дг0_089. Зависимость, аналогичная приведенной, существует между средней массой горбуши Пороная и суммарной величиной вылова основных стад горбуши, нагуливающихся совместно с поронайской в водах Курило-Камчатского района. Для нечетных лету = 1853 r^0_099; для четных — у = 1619 дг0'1. Поскольку уменьшение размеров поронайской горбуши наблюдалось одновременно с увеличением численности рыб в местах нагула, оно, скорее всего, и является следствием увеличения численности. Трудно сказать, что именно явилось непосредственной причиной измельчания. Оно могло быть вызвано ограниченностью кормовой базы, но не исключено, что замедление роста есть следствие повышенной плотности рыб в нагульных скоплениях, оказывающей угнетающее действие на рост через механизмы эндокринной системы. Как бы то ни было, при увеличении численности горбуши, нагуливающейся в Курило-Камчатском районе, она становится мельче. В западном районе Сахалина у горбуши реки Покосная зависимости величины массы от численности поколений не наблюдается (см. рис. 26). В первом случае коэффициент корреляции между массой горбуши и величиной вылова ее у юго-западных районов Сахалина равен 0,031, во втором — 0,158 (0,05 < Р).
Исходя из приведенных выше данных, можно сделать вывод, что уменьшение размеров горбуши, заходящей в реки западных районов Сахалина, обусловлено селективным воздействием японского морского промысла, а уменьшение размеров горбуши в восточных районах Сахалина связано, в первую очередь, с увеличением численности стад, использующих общие с ней нагульные участки в море, и, в меньшей степени, — с селективностью морского промысла.