Воспроизводство горбуши
Условиям развития икры и личинок тихоокеанских лососей в грунте нерестилищ посвящена обширная литература как у нас в стране, так и за рубежом. В настоящее время подробно изучены гидрологический, термический и гидрохимический режимы нерестилищ практически всех видов лососей, а также требования различных видов лососей и их внутривидовых форм к среде на разных этапах онтогенеза [Леванидов, 1969; Смирнов, 1975].
Поскольку в течение эмбрионально-личиночного периода жизни лососей основными элементами их физического окружения являются грунт нерестовых гнезд и омывающая эмбрионы вода, то от качества последних зависит успешность эмбриогенеза. Выживание эмбрионов обеспечивается неподвижностью грунта, его проницаемостью для воды, малым содержанием ила и, в первую очередь, органического ила, на окисление которого расходуется большое количество растворенного в воде кислорода. Вода, омывающая эмбрионы, должна содержать достаточное количество кислорода, расход ее должен быть достаточно высок, чтобы обеспечить как доставку эмбрионам кислорода, так и удаление продуктов их выделения.
Качество грунта и вод подруслового потока даже в пределах одного нерестилища изменяется из года в год. Оно зависит от особенностей уровневого режима, с которым связаны заиляемость грунта и его стабильность, нарушаемая во время паводков, от водности года, определяющей расходы подруслового потока и уровень грунтовых вод. Условия существования эмбрионов в грунте во многом определяются жизнедеятельностью самих лососей, так как производители при устройстве гнезд проделывают огромную механическую работу по перекапыванию грунта, существенно мелиорируя нерестилище, освобождая грунт от ила, делая его более рыхлым и проницаемым для воды. В связи со сказанным выше понятно то внимание, которое уделяется исследователями лососей влиянию на эффективность нереста расходов воды и плотности заполнения нерестилищ производителями.
Установлено, что в реках материкового побережья Охотского моря и в бассейне Амура значительное количество эмбрионов горбуши и летней кеты гибнет во время зимнего падения уровня воды от промерзания нерестилищ [Смирнов, 1947; Костарев, 1964а; Енютина, 1972]. В то же время, на Сахалине промерзание нерестилищ незначительно изза относительно стабильного уровня рек в зимний период и обильных снегопадов [Канидьев, 1967; Воловик, Ландышевская, 1968].
Отрицательное влияние паводков на условия нереста лососей отмечалось неоднократно [Костарев, 19646; Гриценко, 1967; Воловик, Ландышевская, 1968; Чупахин, 1975; Davidson, Hutchinson, 1943; Pritchard, 1948; Neave, 1952; Wickett, 1958; McNeil, 1966; 1969]. Рыба, нерестящаяся во время паводков, откладывает в бугры меньше икры, так как значительная часть ее уносится течением. Подвижки грунта вызывают гибель икры, находящейся в буграх.
Исследователи воспроизводства лососей единодушны в оценке влияния увеличения численности производителей на эффективность нереста одной (средней) самки. При увеличении количества нерестящихся рыб на единицу площади нерестилищ среднее количество икры, откладываемой в бугор, уменьшается, так как при высокой плотности заполнения производители мешают нересту друг друга, повторно перекапывают ранее устроенные бугры, что приводит к увеличению потерь икры [Леванидов, 1969; Pritchard, 1948; Neave, 1953; Wickett, 1958; Hunter, 1959].
Мнения относительно влияния плотности заполнения нерестилищ на численность молоди, продуцируемой всем нерестовым стадом, расходятся. Одни исследователи приходят к выводу, что увеличение численности производителей в конечном счете всегда приводит к увеличению количества молоди, выходящей из грунта, компенсируя снижение эффективности нереста одной самки [Леванидов, 1969; Pritchard, 1948], другие получают результаты, показывающие, что увеличение численности производителей до какого-то предела сопровождается увеличением суммарного выхода молоди, но превышение этого предела приводит к его снижению [Wickett, 1958; Hunter, 1959]. У. Уиккет [Wickett, 1958], В.Н. Иванков и В.Л. Андреев [1972] считают, что оптимальная величина заполнения нерестилищ, обеспечивающая наибольший выход молоди, должна определяться эмпирически для каждого случая, так как в разных водоемах она различна.
Мы поставили целью выяснить, каковы региональные особенности выживания горбуши в период пребывания ее икры и личинок в грунте, как зависит эффективность нереста от средней абсолютной плодовитости, расходов воды и численности производителей. При этом особое внимание уделялось факторам, которые в принципе могут контролироваться человеком, а также факторам, имеющим прогностическую ценность.
По времени нерест горбуши Сахалина совпадает с летней меженью. Это обстоятельство в дальнейшем в значительной мере предохраняет бугры горбуши от промерзания, так как при зимнем падении уровня подавляющее их большинство остается под водой. Обычно в августе уровень воды в реках Сахалина относительно постоянен. Паводки, вызываемые дождями, отмечаются довольно редко и бывают гораздо слабее сентябрьских паводков, тогда как последние подчас значительно превосходят весенние. В течение зимних месяцев уровень воды на нерестилищах горбуши весьма стабилен.
Средняя августовская температура воды на нерестилищах горбуши колеблется от 12 до 16° С. От начала к концу нереста происходит снижение температуры с 20-21 до 1011 °С в теплые годы и с 16-17 до 6-8° С в холодные. Температура воды на нерестилищах опускается в ноябре практически до нуля и лишь в апреле поднимается до 0,5° С. Обильные снегопады и стабильный уровень воды хорошо защищают гнезда горбуши от промерзания, которое наблюдается на Сахалине редко и в небольшом объеме.
На Сахалине горбуша откладывает в бугры от нескольких десятков до полутора тысяч икринок. Среднемноголетнее количество икры, откладываемой горбушей в разных реках, различается несущественно и составляет около 400 шт. (табл. 12). Икра, отложенная в бугры, составляет 20-30 % от средней абсолютной плодовитости. Следовательно, 70-80 /о икры горбуши теряется при нересте. Из этого количества обычно не более 1 % приходится на икру, оставшуюся в брюшной полости самок, основная же масса гибнет, будучи унесенной течением. Через 1-2 мес. после нереста средний отход эмбрионов в буграх обычно не превышает 10 % и лишь изредка достигает 15-25 % (см. табл. 12). К февралю — марту происходит снижение числа эмбрионов в буграх за счет того, что часть погибших икринок полностью разлагается или поедается беспозвоночными. В среднем за годы наблюдений такое сокращение по разным рекам составляет 6,9-21,7 % от количества икры, учтенной во время осеннего вскрытия бугров. Средние показатели выживания эмбрионов в контролируемых реках удивительно сходны: из одного бугра выходит около 300 личинок. Таким образом, на Сахалине выживаемость горбуши в нерестовых буграх высока. В среднем она составляет 68-82 % от отложенной икры (и лишь в отдельные годы сокращается до 25-50 %), или 17-19 % от средней абсолютной плодовитости.
Пытаясь определить влияние различных факторов на результативность нереста и выживание эмбрионов, мы остановились на горбуше ключа Холодный (бассейна реки Поронай), поскольку она длительное время находится под наблюдением. Результаты анализа приведены на рис. 17-19 и в табл. 13.
Связь между количеством икры, откладываемой в бугры, и средней абсолютной плодовитостью отсутствует. Между численностью производителей, зашедших в ключ Холодный, и средним количеством икры в бугре существует отрицательная линейная зависимость. Подобная же зависимость отмечена между численностью производителей и средним количеством икры в буграх, выраженным в процентах от средней абсолютной плодовитости (см. рис. 17,а).
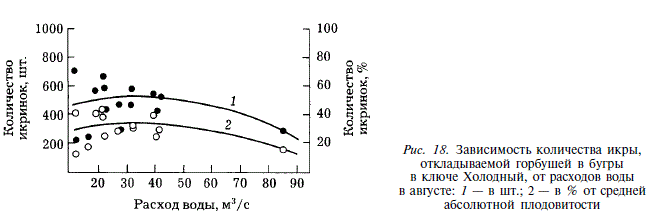
Между расходом воды в августе и количеством отложенной икры (в штуках и в процентах от средней абсолютной плодовитости) существует нелинейная зависимость, описываемая уравнением параболы второй степени (см. рис. 18). Такой характер связи между этими двумя показателями объясняется уменьшением доступной для производителей площади нерестилищ при низких расходах воды, что приводит к повышению плотности их заполнения и, следовательно, к снижению количества икринок, откладываемых в бугор. По мере увеличения расходов воды влияние плотности заполнения нерестилищ производителями ослабевает, и начиная с определенного момента, расход воды становится доминирующим фактором, снижающим количество икры, откладываемой в бугры.
Связи между количеством икры в буграх (в процентах от абсолютной плодовитости) и расходами воды в сентябре не обнаружено. На этом основании можно предположить, что во время паводков в ключе Холодный количество икры в буграх уменьшается за счет выноса ее течением во время нереста, а не за счет размывания уже устроенных бугров. Не обнаружено также связи и между суммой осадков в декабре и январе и количеством живых эмбрионов (в штуках и в процентах от количества отложенной икры), находящихся накануне выхода из грунта. Это подтверждает выводы, что гибель эмбрионов горбуши от промерзания в сахалинских реках невелика (см. табл. 13).

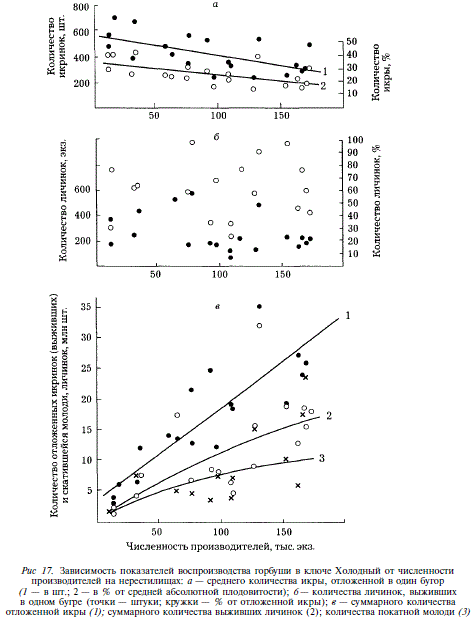
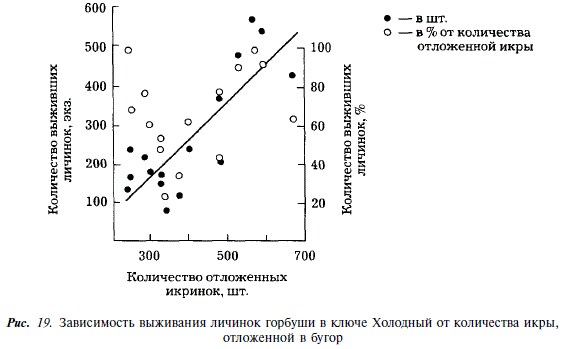
Между количеством икринок, отложенных в бугры, и количеством личинок, находящихся в них накануне выхода, существует положительная линейная зависимость. Связь между количеством отложенных икринок и процентом выживших личинок отсутствует (см. рис. 10). Это дает основание считать, что в условиях ключа Холодный количество отложенной в бугор икры не сказывается на ее дальнейшем выживании. Зависимость между количеством отнерестившихся производителей и процентом выживших личинок не прослеживается. Между количеством отнерестившихся производителей и количеством выживших личинок существует отрицательная корреляция, не достигающая, однако, вероятности 0,95 (см. рис. 17,6). Сказанное относится к результатам нереста каждой самки отдельно.
Суммарное количество икры, откладываемой нерестовым стадом, линейно увеличивается с увеличением численности производителей (см. рис. 17,в, табл. 13). Зависимость суммарного выживания личинок от численности отнерестившихся производителей описывается уравнением степенной функции. Аналогичная зависимость существует между численностью производителей и количеством покатной молоди. Следовательно, увеличение численности производителей горбуши в ключе Холодный сопровождается увеличением суммарного количества выживших личинок и скатившейся молоди.
Наши наблюдения проводились в годы как с низкой, так и с высокой численностью горбуши. В периоды подъема численности плотность заполнения нерестилищ в одном из притоков ключа Холодный — в ключе Березовый — достигала 569 экз. на 100 м, в самом ключе Холодный — 271 экз., что не приводило, однако, к снижению количества покатной молоди. Поэтому, если оптимум заполнения нерестилищ этого водоема и существует, то он выше наблюдавшихся нами величин.
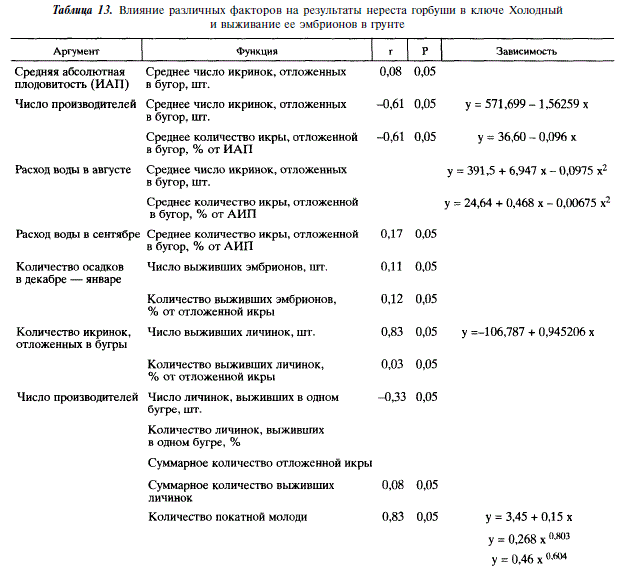
Анализируя зависимость количества покатной молоди от количества отнерестовавших производителей в разных районах Сахалина, мы видим, что в реках Даги, Бахура и Покосная увеличение численности производителей сверх какого-то предела приводит к уменьшению численности покатной молоди (рис. 20, табл. 14). Пределы эти равны соответственно 270; 160 и 290 тыс. экз. производителей, или 99; 460 и 242 особи на 100 м2 нерестилищ. В отличие от этих рек, в Лютоге подобного снижения не наблюдается: максимальная за годы наблюдений плотность заполнения нерестилищ (426 экз. на 100 м2)не привела к снижению количества покатников.
Различия в количестве производителей, обеспечивающих максимальный скат молоди, скорее всего, связаны с особенностями популяционной структуры горбуши разных районов Сахалина. Это количество выше там, где сложнее популяционная структура. Различные экологические требования к условиям нереста и эмбриогенеза у отдельных популяций, видимо, позволяют наиболее эффективно использовать нерестовые водоемы. Хотя следует иметь в виду, что, в свою очередь, возможность их воспроизводства в том или ином водоеме зависит от наличия соответствующих условий. Так или иначе, рассматриваемая ситуация служит дополнительным косвенным подтверждением экологической обособленности популяций горбуши.
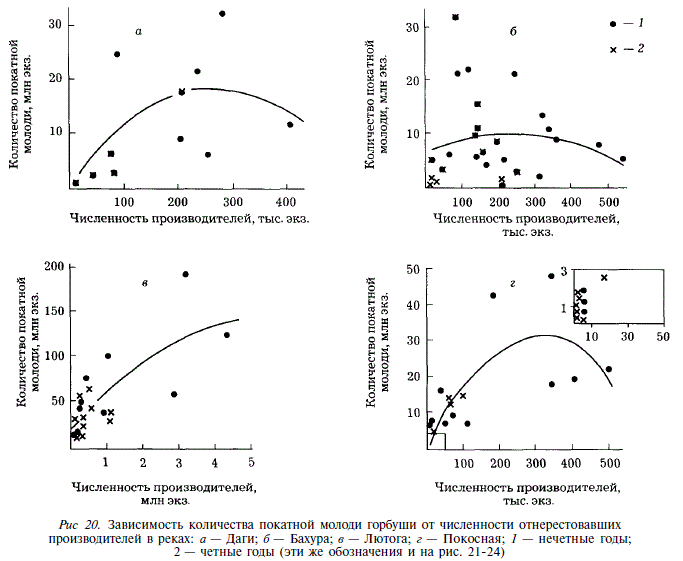
Рассмотрим, как зависит величина возврата рыб от количества покатной молоди. В реках Лютога и Покосная увеличение количества покатников сопровождается увеличением количества вернувшихся рыб (рис. 21, табл. 14). Коэффициент корреляции равен соответственно 0,80 и 0,72. Однако разброс точек на графике столь велик, что использование этой зависимости в прогностических целях неперспективно. Так, в Покосной скат 13-16 млн. мальков обеспечивает возврат от 30 до 200 тыс. взрослых особей.
Иная ситуация наблюдается в реках Бахури и Даги, где увеличение количества покатной молоди лишь до какого-то предела сопровождается увеличением возврата, после чего количество вернувшихся рыб снижается (см. рис. 21, табл. 14). В Даги максимальный возврат обеспечивается скатом 25 млн. мальков, в Бахуре — 17 млн. Подобная ситуация свидетельствует о действии на молодь в прибрежье моря факторов смертности, зависящих от плотности.
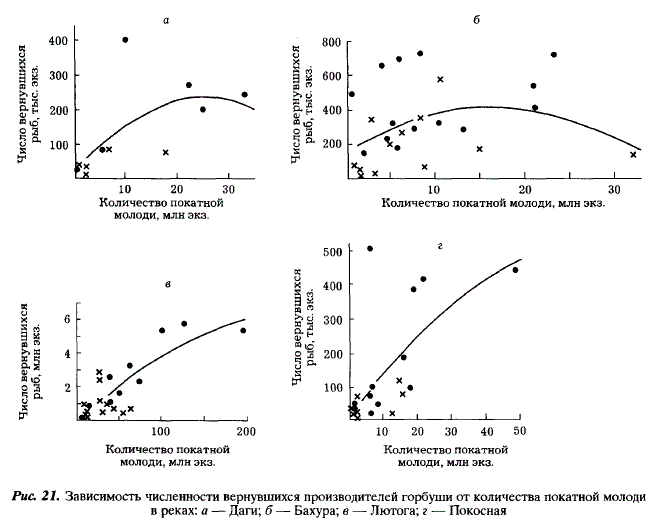
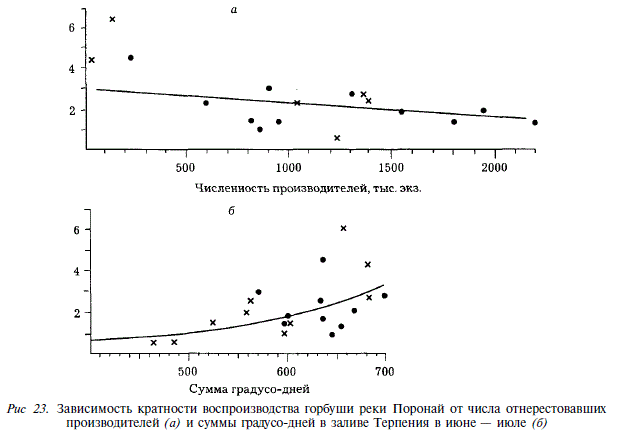
Коэффициенты возврата (Кв) горбуши уменьшаются по мере увеличения количества покатной молоди. Из рис. 22 следует, что факторы смертности, зависящие от плотности, действуют на молодь горбуши не только в реках Бахура и Даги, как это было показано выше, но и в реках Покосная и Лютога. Так, в Покосной увеличение количества покатной молоди с 2 до 50 млн. экз. приводит к пятикратному уменьшению Кв. В Лютоге подобная тенденция прослеживается для поколений четных и нечетных лет в отдельности, хотя в целом для всех лет наблюдения она отсутствует. У горбуши Бахури зависимость Кв от количества покатной молоди становится более четкой, если из анализа исключить годы с резкими отрицательными аномалиями температуры воды в прибрежье моря (1969; 1981; 1983) (см. рис. 22,а), когда влияние на коэффициент возрата термики затушевывает влияние численности покатников. У горбуши реки Поронай, где данные по скату ограничены, мы, тем не менее, обнаруживаем сходный эффект. Кратность воспроизводства уменьшается по мере увеличения количества отнерестовавших рыб (рис. 23), что также является свидетельством действия факторов смертности, зависящих от плотности, хотя рассматриваемая зависимость обусловлена их действием не только в море, но также на нерестилищах.
Ранее было показано, что ряд хищных рыб переходит на питание молодью лососей в период ее интенсивного ската, когда она становится массовой формой макропланктона [Чуриков, Гриценко, 1983]. Естественно, что в годы с высокой численностью покатная молодь привлекает большее количество хищников, а продолжительность периода их воздействия также увеличивается.
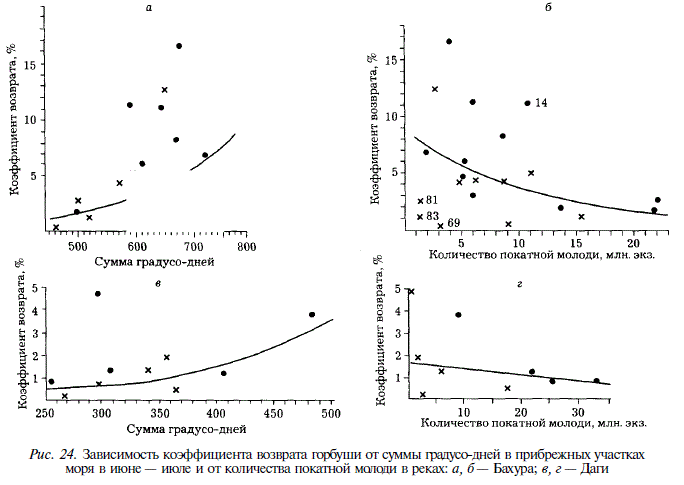
Известно, что смертность животных значительно возрастает при смене среды обитания в процессе онтогенеза. Так, для проходных рыб установлено, что при попадании в море их молодь гибнет от термического шока и от резкого изменения солености. Понижение температуры воды, приводящее к снижению двигательной активности молоди, увеличивает ее доступность для хищников. В годы со значительными отрицательными температурными аномалиями ухудшается обеспеченность молоди пищей, замедляется ее рост, что также не способствует выживаемости. Зависимость урожайности поколений южнокурильской горбуши от термических условий в прибрежье моря убедительно продемонстрирована В.Н. Иванковым с соавторами [1984а]. В этой связи интересно рассмотреть зависимость выживаемости горбуши (в нашем случае характеризуемой Кв) от термических условий в прибрежных участках моря. В качестве показателя последних мы используем температуру поверхностного слоя воды. У горбуши рек Бахура и Лютога обнаружена зависимость К от суммы градусо-дней в июне — июле, описываемая уравнением экспоненциальной функции (рис. 24; см. табл. 14). Аналогичным образом от суммы градусо-дней в июне — июле в заливе Терпения зависит кратность воспроизводства горбуши реки Поронай (см. рис. 13, табл. 14). Это обстоятельство дает нам основание предполагать существование подобной зависимости и для реки Даги, хотя положение эмпирических точек на рис. 24,6 не столь четко характеризует тенденцию, как на рис. 24,а, в и 23,6. При количественной оценке зависимости Кв от суммы градусо-дней в Даги мы исключаем точку с ординатой 4,8, далеко отстоящую от основной группировки точек и нарушающую тенденцию.
Зависимость К от термических условий в море может быть использована в прогностических целях. Как минимум мы сможем предвидеть резкие снижения численности в годы с большими отрицательными аномалиями и увеличения — в годы с большими положительными аномалиями.
У горбуши реки Покосная зависимость Кв от термики прибрежных вод обнаружить не удалось. Отсутствие ее на юго-западном Сахалине свидетельствует об отличии условий формирования поколений от таковых в заливе Анива и на восточном Сахалине. Молодь горбуши, скатывающаяся из рек восточного Сахалина и залива Анива, попадает в открытые участки побережий. Восточносахалинская горбуша попадает в зону действия холодного Сахалинского течения. В этом районе она периодически подвергается воздействию нагонного восточного ветра, приносящего к берегам острова холодные воды и льды Охотского моря. Так, в июне 1969 г. молодь горбуши была перетерта льдами, пригнанными к побережьям залива Терпения и юго-восточного Сахалина. Прибрежные воды югозападного Сахалина защищены от подобных воздействий. Кроме того, там имеются удобные для нагула молоди горбуши в первые дни после ската прогреваемые мелководья, огражденные от моря рифовой грядой, где молодь может успешно адаптироваться к морским условиям.