Популяционная структура горбуши
Показано существование генетических различий между горбушей разных сроков хода [Алтухов и др., 1983], соответствующей сезонным расам, описанным В.Н. Иванковым [1967,1971]. В пределах рас горбуша также оказалась неоднородной, что может служить подтверждением реальности "подходов", описанных СП. Воловиком [1967а].
Новым шагом в понимании популяционной структуры горбуши было обнаружение генетических различий между линиями поколений горбуши четных и нечетных лет. Эти различия касаются частот фенотипов различных ферментных систем [Салменкова и др., 1981; Алтухов и др., 1983; Ермоленко и др., 1983; Гагальчий, 1985, 1986; Aspinwall, 1974], хромосомного полиморфизма [Горшкова, Горшков, 1983], морфометрических и остеологических признаков [Горшков, 1983]. Относительно причин возникновения этих различий высказываются две гипотезы, рассматриваемые авторами как конкурирующие. Согласно одной из них, различия обусловлены селективным преимуществом различных морф в четные и нечетные годы. При этом предполагается существование связи характера полиморфизма с различиями в численности смежных поколений [Горшков и др., 1985]. Согласно второй гипотезе, причиной различий является дрейф генов, который мог сыграть решающую роль в историческом прошлом при малой численности основателей генеративных линий [Салменкова и др., 1981; Омельченко, 1985; Aspinwall, 1974; Beacham, 1985]. Н. Эспинвол и Т. Бичем предполагают, что горбуша четных и нечетных лет переживала последнее оледенение в различных рефугиумах: первая — на севере, в р. Юкон и северной Аляске, вторая — на юге, в р. Колумбия. Гипотезу, объясняющую возникновение различий действием отбора, сторонники второй гипотезы считают несостоятельной, так как, по их мнению, в пределах регионов отбор в многолетнем плане должен действовать одинаково на обе линии.
Здесь уместно сказать о еще одном интересном результате популяционно-генетичсских исследований тихоокеанских лососей, который, к сожалению, практически не учитывался при интерпретации данных в разработке перечисленных выше точек зрения относительно популяционной структуры. Рядом авторов была показана селективная природа полиморфизма некоторых генетических признаков, а также было установлено, что направление селекции может в онтогенезе меняться на противоположное [Кирпичников, 1977,1983; Кирпичников, Иванова, 1977; Кирпичников, Муске, 1981; Кирпичников и др., 1986; Алтухов и др., 1986; Животовский и др., 1986]. Кроме того, было показано, что при возрастании численности лососей в нерестовых стадах увеличиваются частоты редких аллелей [Кирпичников и др., 1986].
В.Т. Омельченко [1985] предполагает, что обнаруженные различия "являются отражением достаточно глубокой адаптированной дифференциации" горбуши четных и нечетных лет. В подтверждение он ссылается на работу СЕ. Дягилева и Н.Б. Маркевича [1979], которые различиями в сроках нерестового хода объясняют различия в выживаемости горбуши по четным и нечетным годам при акклиматизации на европейском севере. Подобное мнение высказывает Т. Бичем [Beacham, 1985] в отношении американской горбуши, ссылаясь на данные У. Риккера [Ricker, 1962] о соотношении численности горбуши четных и нечетных лет в разных широтах.
Популяционная структура горбуши, по мнению В.Н. Иванкова [1986], включает четыре уровня: I — смежные поколения четных и нечетных лет; II — сезонные расы; III — стада (локальные популяции), представляющие собой сложные популяционные системы; IV — демы (менделевские популяции). Несколько отличается от нее схема, предлагаемая В.Т. Омельченко [1985], который также различает четыре уровня генетической дифференциации: I — между поколениями четных и нечетных лет; II — между совокупностями популяций из географически отдаленных районов (Азия, Америка); III — между горбушей разных сроков размножения в одном географическом районе; IV — между субпопуляциями в пределах раннеи позднемигрирующих популяций в пределах одного нерестового хода. Ю.П. Алтухов с соавторами [1983] обнаруживают у горбуши Сахалина I, III и IV уровни приведенной выше иерархии.
В корне отличных взглядов придерживаются М. К. Глубоковский и Л. А. Животовский [1986]. Ими предлагается концепция "флуктуирующего стада" горбуши. По мнению этих авторов, в результате действия миграционных барьеров происходят флуктуации миграционных потоков из мест нагула к местам нереста, вследствие чего в одних районах ареала популяционная структура может временно отвечать концепции локального стада, в других — различным вариантам континуальной модели, либо модели с полным перемешиванием. Сезонные расы горбуши, как считают эти исследователи, образуют независимый от обмена мигрантами и генами уровень внутривидовой дифференциации, возникающей, видимо, в эпигенезе особей каждого поколения. В связи с вышесказанным локальное стадо не может, как утверждают авторы, служить единицей прогнозирования, искусственного воспроизводства и промыслового использования горбуши. К сожалению, в работе не указываются конкретные миграционные барьеры и не приводится примеров их действия. В исследовании популяционной структуры горбуши мы ставим перед собой следующие задачи: 1) выделение наиболее крупных популяционных группировок (т.е. популяций второго иерархического уровня) у вида в целом; 2) выделение наиболее мелких локальных группировок у сахалинской горбуши; 3) анализ природы сезонных рас; 4) разработку гипотезы, объясняющей природу генетических различий между поколениями четных и нечетных лет.
В качестве признаков для выделения популяций второго иерархического уровня мы использовали сроки массового нерестового хода и нереста, места нереста и морского нагула. Особи, принадлежащие к популяциям, выделенным по этим признакам, должны обладать сходной продолжительностью морского нагула и сходными сроками созревания, что, в свою очередь, предполагает их определенное эколого-генетическое единство. Действительно, сходство сроков созревания и нереста должно способствовать выживанию потомства при обмене особями между популяциями более низких рангов. Различия же сроков созревания и продолжительности морского нагула у горбуши, воспроизводящейся в отдельных районах ареала, весьма значительны. Так, если в северной части Берингова моря скат молоди происходит в мае — июне, а нерест — в июле, то в штате Вашингтон скат отмечен в марте — апреле, а нерест — в октябре — ноябре. Следовательно, продолжительность морского нагула в первом случае равна примерно 12, а во втором — 18 мес. У Риккер [Ricker, 1972] сообщает, что у горбуши, выпущенной молодью в реки, весьма удаленные от донорских водоемов, сохранились сроки нерестового хода, свойственные родительскому стаду. Это, по его мнению, свидетельствует о наследственной закрепленности данного признака. Подобное свойство, видимо, характерно для всех лососей. Так, Т. Оказаки [Okazaki, 1982a] описывает его у кеты.
При разделении горбуши азиатского побережья мы использовали литературные данные о сроках захода производителей в реки [Кузнецов, 1928; Миловидова-Дубровская, 1937; Семко, 1939; Агапов, 1941; Кагановский, 1949; Двинин, 1952; Иванков, 1967, 1971; Воловик, 1967а; Енютина, 1972; Пушкарева, 1975; Ishida, 1967] и собственные наблюдения, проведенные на некоторых реках Сахалина. Для горбуши американского побережья более полным оказались данные по срокам нереста, в обобщенном виде приведенные в двух сводках [Atkinson et al., 1967; Aro, Shepard, 1967]. Кроме них, мы использовали результаты многолетних наблюдений за нерестом горбуши в отдельных реках американского побережья, изложенные в ряде работ американских и канадских авторов [Pritchard, 1937; Foerster, Ricker, 1953; Hunter, 1959; Merrell, 1962; Helle, 1970 и др.].
В Северной Пацифике существует четыре района морского нагула горбуши [Бирман, 1967; Hartt, 1962a, b; Ishida, 1962; Kondo et al., 1965; Manzer et al., 1965]: япономорский, включающий акваторию Японского моря севернее 38° с.ш.; камчатско-курильский, ограниченный на севере примерно 52° с.ш., на юге — 38° с.ш., на востоке — 170° в.д.; алеутский, расположенный между 160° в.д., 160° з.д. и 40° с.ш., и восточный, включающий в себя залив Аляска и прилежащие к нему воды океана, ограниченные на юге 40° с.ш. и на востоке 160° з.д.
В япономорском районе нагуливается горбуша, размножающаяся в Амуре, в реках бассейна Японского моря, заливе Анива, реках юго-восточного Сахалина и южных Курильских островов. Из камчатского-курильского района горбуша идет в реки бассейна Охотского моря, а также тихоокеанского побережья острова Хоккайдо. Горбуша из алеутского района нерестится по азиатскому побережью от Чукотки до восточной Камчатки, по американскому — от залива Коцебу до северной границы юго-всточной Аляски, а также на Алеутских островах. Из восточного района горбуша идет на нерест в реки юговосточной Аляски и далее на юг до южной границы ареала.
Нерест горбуши в южных частях ареала происходит в более поздние сроки, чем в северных (табл. 8 и 9). Однако это только тенденция, и строгой закономерности в данном явлении нет. Так, например, в Приморье она нерестится в те же сроки, что в реке Анадырь. Горбуша острова Кодьяк нерестится раньше, чем более северная горбуша реки Кускоквим.
Материалы, приведенные в табл. 8 и 9, свидетельствуют, что сроки хода (или нереста) постоянны на огромных участках ареала. Таких участков мы выделяем девять, и горбушу, воспроизводящуюся на них, предположительно относим к различным популяциям второго ранга (рис. 10). Перейдем к описанию этих популяций.
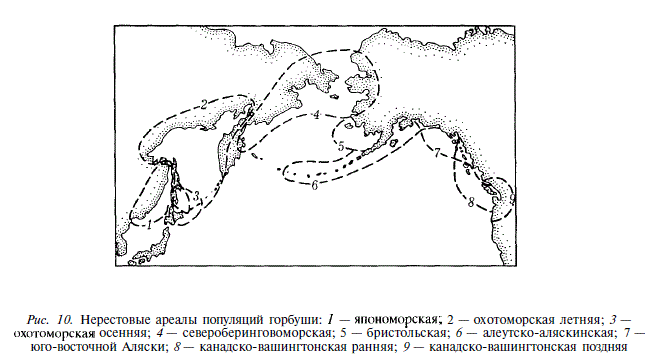

1 . Япономорская . Нерестовый ареал: Приморье, бассейн Амура, западный Сахалин, зал. Анива, юго-восточный Сахалин до 50° с.ш., южные Курильские острова, япономорское побережье острова Хоккайдо. Северная граница распространения по восточному побережью Сахалина установлена мечением нагульной горбуши в Японском море и молоди, выпущенной Айнским рыбоводным заводом [Ландышевская, 1962; Рухлов, Любаева, 1980; Kondo et al., 1965]. О принадлежности к япономорской популяции части горбуши Южных Курил мы судим на основании поимки в Японском море особей, меченных в 1976 г. на рыбоводных заводах острова Итуруп [Рухлов, Любаева, 1980]. Массовый нерестовый ход во II декаде июня — III декаде июля. Нагул в Японском море.
2. Охотоморская летняя . Нерестовый ареал: западная Камчатка, материковое побережье Охотского моря, восточный Сахалин, охотоморское побережье западного Сахалина, зал. Анива, Курильские острова, охотоморское и тихоокеанское побережья острова Хоккайдо. Нерестовый ход с III декады июля по III декаду августа. Нагул в камчатско-курильском районе.
3. Охотоморская осенняя . Нерестовый ареал: восточное побережье острова Сахалин к югу от мыса Терпения, залив Анива, охотоморское (преимущественно) и тихоокеанское побережья острова Хоккайдо, южные Курильские острова. Присутствие этой популяции на Южных Курилах и Хоккайдо наглядно иллюстрируется двумя пиками нерестового хода [Иванков, 1967, 1971; Ishida, 1967]. На Сахалине ее трудно выделить по динамике хода, видимо, из-за относительной малочисленности. О присутствии ее в этом районе мы судим по регулярному октябрьскому нересту, отмечаемому к югу от мыса Терпения, в реках залива Мордвинова и восточного побережья залива Анива, подобно тому, как это имеет место на островах Итуруп и Хоккайдо. К северу от мыса Терпения нерест горбуши в октябре отмечается исключительно редко, видимо, при случайных проникновениях туда части охотоморской осенней популяции. Нерестовый ход с III декады августа по III декаду сентября. Морской нагул в камчатско-курильском районе. Как видим, нерестовый ареал охотоморской осенней горбуши не выходит за пределы южной части ареала охотоморской летней. Тем не менее, мы выделяем ее в качестве популяции второго ранга. Подробнее это выделение мы попытаемся обосновать ниже.
4. Североберинговоморская . Нерестовый ареал: северо-восточная Камчатка, Корякское побережье, река Анадырь, залив Коцебу, река Юкон. Данных о сроках нерестового хода на юго-восточной Камчатке получить не удалось, но, скорее всего, этот район также входит в ареал североберинговоморской популяции. Нерестовый ход в июле. Нагул в алеутском районе.
5. Бристольская . Нерестовый ареал: река Кускоквим, побережье Бристольского залива. Нерест в III декаде августа — сентябре. Нагул в алеутском районе.
6. Алеутско-аляскинская . Нерестовый ареал: п-ов Аляска, Алеутские острова, о. Кодьяк, р. Сустина, залив Кука. Нерест с III декады июля до конца августа. Нагул в алеутском районе.
7. Популяция юго-восточной Аляски . Нерестовый ареал: зал Принца Вильяма, материковое побережье юго-восточной Аляски и Британской Колумбии примерно до устья реки Филлипс (50°20' с.ш.), верхнее течение реки Скина. Нерест в августе и I декаде сентября. Нагул в восточном районе.
8. Канадско-вашингтонская ранняя . Нерестовый ареал: острова, соседствующие с материковым побережьем юго-восточной Аляски и Британской Колумбии, нижняя часть бассейна реки Скина, материковое побережье штата Вашингтон до 48°10' с.ш., верхние участки бассейна реки Фрейзер. Нерест во II декаде сентября — I декаде октября. Нагул в восточном районе.
9. Канадско-вашингтонская поздняя . Северная граница нерестового ареала находится не южнее реки Фрейзер. Однако весьма вероятно, что эта популяция распространена гораздо севернее, вплоть до пролива Саммер. Южная граница нерестового ареала совпадает с южной границей видового ареала горбуши. В сводке Аткинсона с соавторами [Atkinson et al., 1967] в качестве таковой выступает река Пиаллап [с. 88]. Самые южные нерестилища на ней расположены на 47° с.ш. Таким образом, нерестовый ареал этой популяции в значительной мере совпадает с ареалом канадско-вашингтонской ранней. Эта ситуация напоминает совпадение ареалов охотоморской летней и осенней горбуши, однако в данном случае перекрывание неполное. Нерест во П-Ш декадах октября, нагул в восточном районе.
Выделенные нами популяции окажутся реальными при соблюдении одного из двух условий: а) ареалы их не должны совпадать, признаки, по которым они выделяются, должны резко различаться на стыке ареалов; б) если ареалы перекрываются, то в зоне перекрывания должны включаться механизмы, ограничивающие межпопуляционную скрещиваемость. Первое условие, как видим, не соблюдается, так как ареалы популяций перекрываются весьма широко. Посмотрим, в таком случае, как ведут себя популяции в зоне перекрывания. Для этого воспользуемся несколькими примерами, имеющимися в литературе. Так, существуют описания размножения горбуши в зоне перекрывания ареалов популяций юго-восточной Аляски и алеустско-аляскинской [Helle, 1970], канадско-вашингтонской ранней и канадско-вашингтонской поздней [Ward, 1959; Vernon, 1962], популяции юго-восточной Аляски и вашингтонской ранней [Skud, 1958; Sheridan, 1962], охотоморской летней и осенней [Иванков, 1967, 1971]. Во всех случаях наблюдается весьма сходная ситуация: между горбушей разных сроков хода обнаруживается пространственно-временная изоляция. В пределах бассейна одной реки используются разные нерестовые участки, там же, где нерест происходит на одном и том же нерестилище, оно используется в разные сроки. Смешиваемость в последнем случае невелика, так как рыбы конца раннего и начала позднего хода пребывают в разной готовности к нересту.
Некоторое основание для суждения о стабильности границ популяционных нерестовых ареалов дают наблюдения, проведенные на реке Тымь, где воспроизводится охотоморская летняя горбуша. А.Я. Таранцом в 1933 г. и мною в 1968 г. отмечался заход горбуши в эту реку в сроки, характерные для охотоморской осенней популяции (сентябрь — октябрь) [Таранец, 1937]. Эти факты позволяют предположить, что иногда часть особей той или иной популяции заходит на нерестилища, расположенные за пределами нерестового ареала, однако нерест в таких случаях, видимо, не эффективен, так как подобные заходы редки и нерегулярны.
Все изложенное позволяет высказать ряд соображений относительно природы сезонных рас у горбуши. Данные о сроках хода и нереста не дают оснований считать деление этого вида на летнюю и осеннюю формы универсальным свойством и рассматривать эти формы в качестве определенного иерархического уровня. Как мы показали выше, реально существующая ситуация оказалась гораздо сложнее: как на американском, так и на азиатском побережьях обитают не две, а несколько форм горбуши, различающихся сроками хода. Так, популяция юго-восточной Аляски является осенней по отношению к более северной, алеутско-аляскинской, популяции и в то же время летней по отношению к более южной, канадско-вашингтонской ранней. Можно было бы привести и другие подобные примеры. Поэтому мы считаем, что у горбуши нет сезонных рас в классическом понимании этого определения, однако популяции второго ранга в районе перекрывания ареалов ведут себя по отношению друг к другу как сезонные расы. Рассматривая с этой точки зрения осеннюю расу горбуши, описанную В.Н. Иванковым [1967] для южной части Охотского моря, мы оцениваем ее как популяцию второго ранга, сохранившуюся как реликт времени с более холодным климатом в зоне перекрывания ареалов с соседней северной популяцией.
Показательно, что во многом сходная ситуация существует на американском побережье, где ареал самой южной популяции — канадско-вашингтонской поздней — почти полностью умещается в пределах ареала канадско-вашингтонской ранней. Очевидно, что в периоды похолодания климата эти южные популяции распространяются дальше на юг по сравнению с существующими на сегодняшний день границами.
Отсутствие различий по исследованным генетическим признакам у горбуши из разных рек на обширных участках побережья является веским, но все же недостаточным основанием для вывода о ее популяционной неподразделенности, так как сходство признаков может быть обусловлено селекцией. Для выяснения популяционной структуры сахалинской горбуши мы использовали данные СахТИНРО по авиаучету производителей горбуши в сахалинских реках и данные Сахалинрыбвода по величине нерестовых площадей. На Сахалине результаты авиаучета корректируются данными визуального учета, проводимого при пеших обходах нерестилищ. Последнее бывает особо необходимым при экстремально высокой плотности заполнения нерестилищ. Нами рассматриваются реки северовосточного Сахалина (от реки Вал до мыса Терпения), юго-восточного Сахалина (от реки Мануй до реки Долинка) и реки залива Анива. В границах этих участков наблюдается сравнительно большое однообразие природных условий воспроизводства горбуши. Между собой участки существенно различаются характером хода многолетних изменений ее численности. Наиболее крупные реки этих участков — Поронай, Тымь, Найба, Лютога, не включались в анализ.
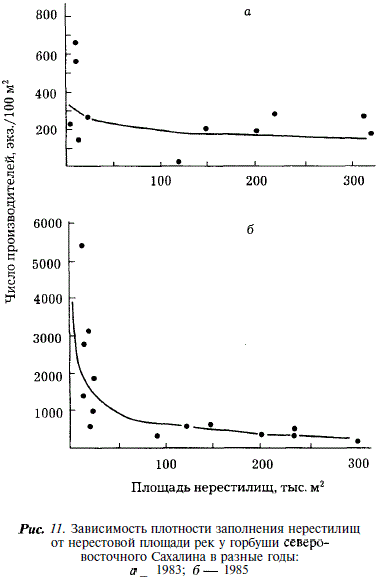
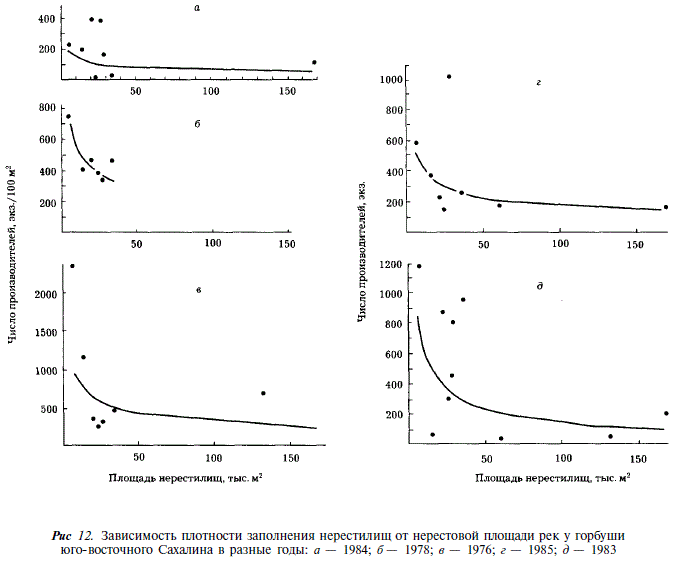
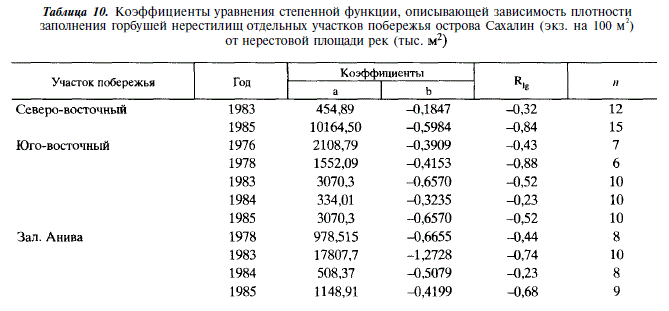
Учет производителей горбуши в реках Сахалина показал, что в отдельных случаях плотность заполнения нерестилищ бывает крайне высока. Так в реках северо-восточного побережья максимальная плотность заполнения нерестилищ достигала 5390 экз/100 м , в реках юго-западного побережья —1180, в реках зал. Анива — 450 шт. на 100 м. При этом наибольшая плотность отмечается в малых реках, площадь нерестилищ в которых не превышает 20 тыс. м. Более того, как видно из рис. 11-13, в ряду рек, ранжированных по величине площади нерестилищ, отмечается закономерное увеличение плотности заполнения по мере уменьшения площади нерестилищ. Наиболее четко эта закономерность проявляется в годы с экстремально высокой численностью горбуши (рис. 11,6; 12,г,д; 13,а,в). В этих случаях связь между площадью нерестилищ и плотностью их заполнения описывается уравнением степенной функции:
(табл. 10). Для удобства сравнения мы пользуемся этим уравнением и в других случаях, где эта зависимость проявляется менее четко.
Первое, что можно предположить для объяснения обнаруженного феномена, — это большая эффективность воспроизводства горбуши в реках с малой площадью нерестилищ. Если такое предположение верно, то мы неизбежно должны допустить, что обмен особями между реками настолько мал, что не может исказить или сгладить результаты различий в условиях воспроизводства, т.е. в каждой реке существует свое стадо горбуши.
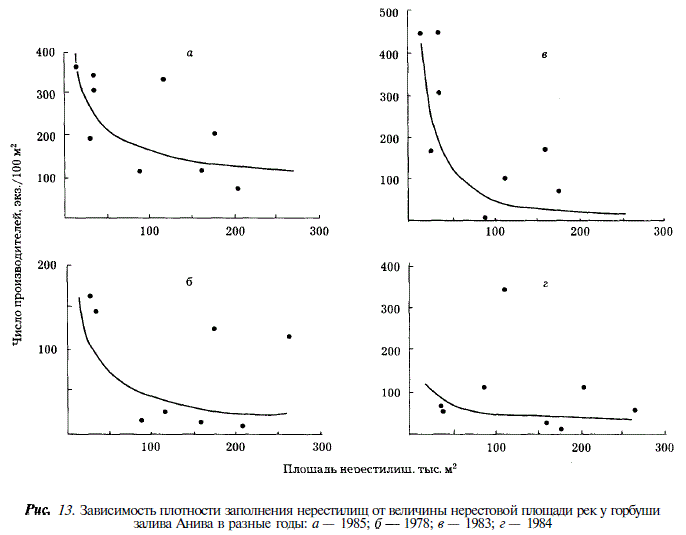
Поскольку реки, анализируемые в пределах отдельных участков, находятся в одинаковых климатических условиях (протекают по районам острова с одинаковым геологическим строением и обладают вследствие этого сходными химизмом вод, морфологией русла, составом грунта нерестилищ, а также гидрологическим режимом), единственным очевидным несходством между ними является разница в нерестовой площади. Сам по себе этот факт на основании имеющихся на сегодня знаний о горбуше не предполагает различий в эффективности ее воспроизводства. Вместе с тем установлено, что превышение плотности заполнения нерестилищ лососями сверх определенной величины приводит к снижению количества покатной молоди. Для сахалинской горбуши эта величина неодинакова в разных реках. Так, на юго-восточном побережье в реке Бахура она равна, 692 экз/100 м в реке Даги (северо-восточное побережье) — 99, в реке Покосная (югозападное побережье) — 119 экз. на 100 м [Гриценко и др., 1989]. Многократное превышение этих величин в малых реках должно было привести к снижению количества покатной молоди и, как следствие этого, к уменьшению возврата. Однако, на юго-восточном Сахалине с 1981 по 1989 г. отмечалась исключительно высокая численность горбуши. Таким образом, объяснение устойчиво наблюдаемого многократного переполнения нерестилищ в малых реках особо благоприятными условиями воспроизводства горбуши в них противоречит данным об оптимальной плотности заполнения нерестилищ. Тем самым популяционная самостоятельность горбуши малых рек закономерно ставится под сомнение и заставляет искать другое объяснение явлению.
В этой связи уместно вспомнить, что горбуша в годы высокой численности появляется в водоемах, где ее до этого не было долгое время. Подобное явление описано в бассейне реки Фрейзер [Vernon, 1962]. А.Н. Канидьев [1968] отмечал восстановление воспроизводства горбуши в ряде сахалинских рек после полного уничтожения ее лесосплавом. Наличие обмена особями горбуши между сахалинскими реками установлено посредством мечения [Рухлов, Любаева, 1980]. Эти факты служат основанием для другого объяснения: в пределах выделенных районов увеличение плотности заполнения нерестилищ горбуши в малых реках происходит вследствие большого количества мигрантов из более крупных рек. При этом необходимо сделать три следующих допущения: 1) эффективность воспроизводства горбуши в реках с различной площадью нерестилищ одинакова при одинаковой плотности их заполнения, хотя строгое равенство необязательно; 2) вероятность захода особи в реку не зависит от размеров реки; 3) процент особей, вернувшихся в родную реку и миновавших ее, одинаков в реках с различной площадью нерестилищ; строгое равенство в данном случае также необязательно, поскольку относительно меньший процент эмигрантов горбуши из большой реки может по абсолютной численности существенно превосходить потомство, вернувшееся в малую реку.
На основании установленных зависимостей мы рассчитали долю иммигрантов в условных реках с разной нерестовой площадью. Эта доля вычислялась как превышение плотностью заполнения нерестилищ в той или иной конкретной реке плотности заполнения нерестилищ наиболее крупной реки участка, принятой в данном районе за точку отсчета, выраженное в процентах от плотности заполнения в первой. При этом предполагалось, что в наиболее крупной реке влияние иммигрантов минимально и им можно пренебречь. На северо-востоке в качестве таковой рассматривалась река Нерпичья (300 тыс. м2 нерестилищ), на юго-востоке — река Фирсовка (168 тыс. м2), в заливе Анива — река Тараннай (263 тыс. м2).
Как следует из данных табл. 11, в годы с высокой численностью горбуши (1983,1985) в реках с нерестовой площадью от 10 до 50 тыс. м2 доля иммигрантов составляет от 28 до 97 %; в реках с площадью нерестилищ 100-200 тыс. м2 — от 7 до 70 %. Однако и в годы с низкой численностью малые реки обычно более чем на половину заполняются иммигрантами. Поэтому есть основание утверждать, что в малых сахалинских реках, таких как Анна, Сима, Игривая, Шешкевича, Пильнги, Пурш-Пурш, Венгери, Б. Хузи, Герань, нет самостоятельных популяций горбуши. Едва ли приходится предполагать существование популяций и в более крупных реках, так как в течение нескольких лет в них должна происходить полная замена особей-автохтонов иммигрантами.
Особо следует остановиться на горбуше крупнейших сахалинских рек: Поронай, Тымь, Найба, Лютога. В эти реки ее производители входят с менее зрелыми половыми продуктами, чем в те сравнительно небольшие реки, которыми мы оперировали в своем анализе. СП. Воловик и А.Е. Ландышевская [1968] пришли к выводу, что различия в степени зрелости производителей сахалинской кеты, наблюдаемые в разных реках во время захода, наследственно обусловлены. Дальнейшие исследования, проведенные на кете острова Хоккайдо, подтвердили подобную точку зрения [Hiroi, 1985]. Это дает основание предполагать, что и горбуша, входящая в крупные реки с менее зрелыми половыми продуктами, чем у рыб, идущих на нерест в малые реки, является репродуктивно обособленной. Поэтому горбушу, входящую в реки Поронай, Тымь, Найба и Лютога, следует рассматривать как обладающую популяционным статусом. Репродуктивный успех при обмене особями между малыми и большими реками должен быть пониженным по сравнению с обменом между только малыми или только большими реками. В то же время обмен особями между большими реками должен быть затруднен из-за значительных расстояний между их устьями, достигающих нескольких сотен километров, в то время как для малых, устья которых отстоят друг от друга на несколько километров, расстояние не является препятствием для обмена особями. В связи с этим при попытках определения межпопуляционных границ уместно вспомнить, что при акклиматизации в Великих озерах горбуша распространялась по побережью на 30-70 км за жизнь одного поколения [Kwain, Lowrie, 1981].
Нам представляется, что у сахалинской горбуши в качестве межпопуляционной границы может рассматриваться полуостров Терпения, где на участке побережья протяженностью более 100 км отсутствуют нерестовые реки. Подобной границей, видимо, является участок юго-восточного побережья, от мыса Свободный до мыса Анива, а также полуостров Шмидта. В этих трех случаях должна происходить изоляция расстоянием. Кроме того, вероятно, существуют границы другой физической природы, представления о которых дают результаты мечения и наблюдения за сроками нерестового хода [Воловик, 1967а; Рухлов, Любаева, 1980; Takagi et al., 1981].
Таким образом, особенностью популяционной структуры сахалинской горбуши является мозаичность расположения нерестовых ареалов популяций низшего иерархического уровня, при которой в ареал популяции, воспроизводящейся во множестве малых рек, вклинивается ареал популяции большой реки. Например, популяция горбуши реки Тымь вклинивается в ареал популяции других рек северо-восточного Сахалина. В целом, нам представляется, что сахалинской горбуше свойственна четырехуровневая популяционная структура, которую мы приводим на рис. 14.
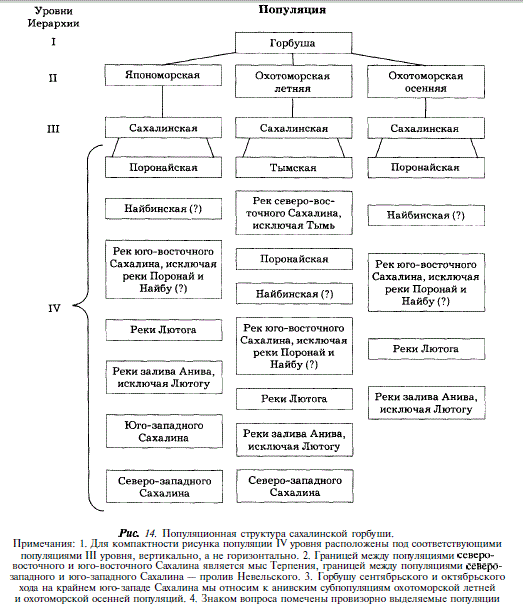
Популяционная подразделенность сахалинской горбуши находит отражение в закономерных изменениях биологических показателей производителей (в частности, длины тела) на протяжении нерестового хода. Для того чтобы понять, как интерпретировать эти изменения в реке, где популяционная структура горбуши сложна, целесообразно рассмотреть их сначала на примере водоема, в котором вероятность популяционной подразделенности воспроизводящейся горбуши минимальна. В нашем материале этому условию наиболее удовлетворяет горбуша Первой Речки, где воспроизводятся представители только япономорской популяции, малые же размеры реки и слабая разветвленность русла не дают оснований предполагать, что горбуша в ней подразделяется локально. Как видно из рис. 15, в Первой Речке размеры производителей увеличиваются от начала к концу хода, либо, что бывает реже, остаются без изменений. На этом основании мы думаем, что подобный характер изменений свойственен популяциям горбуши низшего иерархического уровня в целом.
В свете этих представлений рассмотрим изменения размеров горбуши во время хода в северной части залива Терпения, где вылавливают представителей япономорской, охотоморской летней и осенней популяций, идущих на нерест преимущественно в Поронай. В те годы, когда ход горбуши охвачен наблюдениями от начала июня до конца августа, удается проследить последовательность прохождения различных популяций через зону ставных неводов, что находит отражение в изменении размеров рыб. В течение июня, когда к берегам подходит япономорская горбуша, наблюдается увеличение ее размеров (рис. 16).
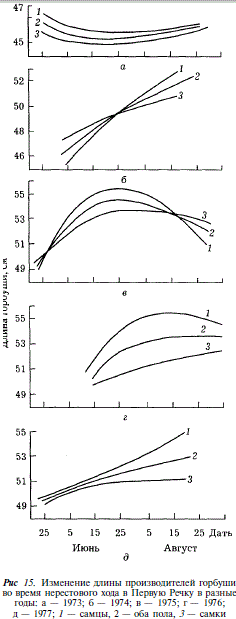
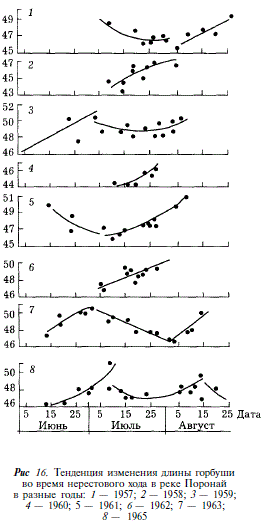
В конце июня — начале июля, по мере того как в уловах уменьшается доля крупных особей, завершающих ход япономорской популяции, и возрастает доля мелких рыб, открывающих ход охотоморской летней горбуши, средняя длина рыб постепенно уменьшается. В дальнейшем, в июле и августе, когда уловы представлены в основном охотоморской летней горбушей, вновь наблюдается увеличение длины рыб, аналогичное тому, какое наблюдалось в процессе хода япономорской горбуши. Прекращение наблюдений в середине августа не позволило нам проследить за ходом охотоморской осенней горбуши. Однако появление в 1965 г. 14 августа наряду с крупной горбушей, длиной 49,2 см, мелкой, длиной 46,9 см, а в дальнейшем появление рыб средней длиной 48,1 см следует рассматривать как наметившуюся тенденцию к уменьшению размеров, связанную с подходом охотоморской осенней горбуши. В те годы, когда наблюдения за ходом горбуши были непродолжительны (1958, 1960, 1962 гг.), по результатам биологических анализов прослеживаются подход только охотоморской летней горбуши и увеличение средних размеров рыб в уловах от начала к концу хода.
Р.И. Енютина [1972], анализируя изменения размеров амурской горбуши в процессе хода, не усматривает в них закономерности. Однако, по ее данным, на протяжении хода можно выделить периоды продолжительностью 10-20 сут, в которые рыба в уловах практически не различается по средней длине. Такая стабильность, видимо, свидетельствует о подходе различных (репродуктивных или нагульных) группировок горбуши.
Рассмотрим вопрос о природе различий генетических признаков между генеративными линиями горбуши четных и нечетных лет. Естественно предположить, что в начальные этапы истории горбуши линий поколений не было и созревание ее происходило в разном возрасте (0 +; 1 +, 2 +), что изредка наблюдается в настоящее время. Видимо, в один из периодов с неблагоприятными условиями воспроизводства отбор стал действовать против мелких, рано созревающих особей, и горбуше стало свойственно созревание практически только в возрасте 1 +. Рассматривая современное воспроизводство горбуши на всем ее ареале, не трудно представить, что уменьшение размеров производителей ниже какого-то определенного предела будет приводить к неспособности устраивать нормальные нерестовые гнезда и, следовательно, к неспособности оставлять потомство. Если в настоящее время можно лишь с большими натяжками представить, как отбор действует против крупных особей и особей в возрасте 2 +, то действие его против мелких особей представляется очевидным. Логично также предположить, что разделение горбуши на две генеративные линии совпало с резким снижением ее численности и имело место на ограниченной акватории водоема происхождения, а современный ареал осваивался обеими линиями одновременно, параллельно и в известной мере самостоятельно.
Возвращаясь к вопросу о причинах генетических различий генеративных линий горбуши, следует отметить, что гипотезы, объясняющие эти различия случайным дрейфом нейтральных мутаций или действием отбора, нам не представляются взаимоисключающими. Так, наблюдаемый в четные годы на всем ареале мономорфизм по локусу фосфоглюкомутазы-1 (изменчивому в поколениях нечетных лет) вполне может быть объяснен действием эффекта основателя. Различия же по другим признакам скорее следует объяснить действием отбора. Так, на примере нерки показано, что распределение частот аллелей локуса лактатдегидрогеназы-В1 изменяется под действием различной температуры, кислорода, наличия продуктов обмена и других меняющихся условий среды. В частности, особое значение придается действию температуры. Носители "северного" аллеля хуже переносят повышение температуры, чем носители "южного" аллеля, и наоборот [Кирпичников, Муске, 1981]. Наглядное подтверждение действия переменных факторов можно получить, если рассмотреть данные Е.А. Салменковой с соавторами [1981] по изменчивости частот фенотипов и генов, маркируемых разными белками, на протяжении нерестового хода горбуши в реке Очепуха (Лесная), которая изучалась методом электрофореза с 1974 по 1979 г. Из этих лет 1977 год отличался аномальностью условий нереста, который происходит при исключительно низком уровне и малых расходах воды, а также при весьма высокой численности производителей, что обусловило чрезмерную плотность заполнения ими нерестилищ. Эмбрионы этого поколения должны были развиваться в условиях пониженного содержания кислорода и повышенной концентрации продуктов метаболизма. Вода в реках имела сильный запах разлагающихся трупов производителей, которые не уносились течением с нерестилищ, как это бывает обычно. Инкубация началась при повышенной температуре воды и продолжалась при резко наступившем охлаждении. Промерзание нерестилищ было выше среднемноголетней величины. В результате в 1979 г. численность дочернего поколения снизилась по сравнению с численностью родительского. Горбуша в 1979 г. характеризуется минимальной изменчивостью всех исследованных признаков на протяжении нерестового хода по сравнению с другими годами [Салменкова и др., 1981, рис. 2]. Видимо, в аномальных условиях 1977 г. смогли выжить особи с узким диапазоном значений признаков, оказавшиеся наиболее приспособленными. Следует вспомнить, что в годы увеличения численности, когда естественно предположить уменьшение давления отбора, увеличивается число особей, носящих редкие аллели [Кирпичников и др., 1986].
Таким образом, принципиальная возможность изменения генетических характеристик популяций горбуши в двух смежных поколениях под действием варьирующих условий среды едва ли может вызвать возражения. Однако противниками гипотезы, объясняющей различия горбуши четных и нечетных лет отбором, ставится под сомнение возможность многолетних различий в действии отбора по четным и нечетным годам. Обратимся в этой связи к следующим фактам. И.Б. Бирман [1985] приводит много примеров двухлетней периодичности природных явлений, которые могут обусловить закономерные различия в урожайности поколений горбуши (колебания речного стока, количества осадков, циркуляции атмосферы). Если обратиться к ситуации последних лет, то можно заметить, например, что у горбуши юго-восточного Сахалина с 1980 г. поколения четных лет формировались при более низкой температуре воды в начале морского нагула молоди, чем поколения нечетных лет. Более того, в 1981 и 1983 гг. их формирование происходило в условиях резких отрицательных аномалий.
Годы 1980 1982 1984 1981 1983 1985 Сумма градусо-дней в поверхностном слое воды в июне — июле 627 647 652 508 526 627
В результате в четные годы наблюдалось прогрессивное снижение численности, а в нечетные годы — увеличение. Согласно упомянутым выше работам B.C. Кирпичникова и соавторов, а также данным С.С. Шварца [1969], полученным на разных группах животных, подобная ситуация должна привести к различиям в генетической структуре поколений четных и нечетных лет. У первых следует ожидать уменьшения генетического разнообразия и увеличения доли "северных", холодостойких, аллелей, у вторых — возрастания генетического разнообразия и доли "южных" аллелей. Таким образом, наблюдаемые генетические различия между поколениями горбуши четных и нечетных лет обусловлены как изначальными различиями, имевшимися у основателей генеративных линий, так и действием отбора в настоящее время.
Наличие двух линий поколений у горбуши нам представляется явлением, преходящим в истории этого вида. Подобные явления должны были возникать в истории многих короткоцикловых видов. Ниже мы встретимся, например, с подобным возрастным составом у корюшки Hypomesus nipponensis.
Со временем, видимо, должно произойти перемешивание линий за счет появления особей, созревающих в более старшем возрасте. Материал для отбора имеется, а время предоставит ситуации, при которых усложнение возрастной структуры окажется выгодным. Пример тому — возникновение второй генеративной линии у горбуши в Великих Озерах [Kwain, Chappel, 1978; Kwain, Lowrie, 1981]. Вместе с тем, генетические различия, подобные обнаруженным между линиями поколений горбуши, могут рассматриваться как предпосылка видообразования. По мере появления в каждой из линий особей, созревающих в возрасте ином, чем 1+, скрещивание их с особями другой линии может не происходить вследствие генетических различий. В этом случае первоначальное разделение вида на две линии поколений, созревающих в одном возрасте, может завершиться возникновением репродуктивно изолированных многовозрастных группировок.
В заключение раздела хочу сказать, что принимаемое многими авторами за основу популяционного деления горбуши существование генеративных линий четных и нечетных лет представляется оправданным. Однако в своей схеме я исхожу из локального деления, рассматривая поколения четных и нечетных лет как возрастные группы локальных популяций, при этом полностью отдавая себе отчет в том, что они сами по себе являются популяциями и требуют соответствующего подхода при ведении промысла и организации воспроизводства.
Что же касается концепции флюктуирующих популяций, то применение ее правомочно к популяциям низшего иерархического уровня, наименее устойчивым во времени, и не только у горбуши, но и у множества других видов животных. Разница между горбушей и другими лососями не в сути явления, а в его размерах. Если у нерки флюктуирующие популяции могут быть представлены несколькими сотнями или тысячами особей и занимать в период нереста весьма небольшую площадь, то у горбуши их численность может достигать нескольких миллионов особей, а нерестовый ареал занимать реки, впадающие на протяжении нескольких сотен километров побережья. Соответственно, действие минимальной промысловой организации (например, бригады рыбаков) в первом случае базируется на множестве флюктуирующих популяций, во втором — одна флюктуирующая популяция способна обеспечить деятельность множества промысловых организаций.
